Биофизика және термодинамика: тірі жүйелердегі энергия түрленуі мен қайтымсыз процестер

Биофизика және оның мәселелері. Термодинамика»
1. Биофизикажәне оның мәселелері.
Биофизика- әртүрлі сатыдағы биологиялық жүйелерде болып жатқан
физикалық және химиялық процестерді зерттейтін ғылым. Биофизиканың зерттейтін объектісі биологиялық материалдар, яғни тірі организмдер. Олай болса физиканың өлі табиғат үшін ашылған заңдарын өзгеріссіз тірі организмге қолдануға болмайды. Оның себебі тірі организм - биологиялық жүйе, үнемі динамикалық қозғалыста болады және гетерогендік әртекті жүйе болып саналады. Биофизиканың ғылым болып қалыптасуына физика, химия, физиология, математика, биохимия тәрізді ғылымдардың зор ықпалы тиді. Осы ғылымдардың негізінде дүниеге келген биофизиканың өз заңдылықтары, өз әдістері бар. Биофизиканың мынадай салалары бар: 1) молекула биофизикасы, ол организмді түзетін биологиялық молекулалардың құрылысы мен физикалық қасиеттерін қарастырады. Сол сияқты биологиялық процестердің кинетикасы мен термодинамикасын қарастырады; 2) клетка биофизикасы. Бұл сала клетканың ультрақұрылысын, оның физикалық және химиялық ерекшеліктерін және клетканың өтімділігін, биологиялық потенциалын қарастырады; 3) күрделі жүйе биофизикасы (немесе басқару және реттеу процесінің биофизикасы) . Сонымен қатар, биофизика организмге деген физикалық факторлар әсерін, иондалған сәуленің биологиялық әсерін (рабиобиология), көз оптикасын, қозғалыс, тыныс алу, иіс сезу, есіту, қан айналыс органдарының жұмыс әрекетін қамтиды. Биофизика ғылым ретінде бүгінгі күн кең дами бастады.
2 Термодинамиканың негізгі түсініктері.
Термодинамика физиканың материя қозғалысының жылулық формасын және сонымен байланысты физикалық құбылыстарды зерттейтін бөлімі. Термодинамикадағы басты орында тұрған ол энергия мен жұмыс арасындағы қатынас. Термодинамикалық жүйе деп өзара бір-бірімен және басқа денелермен зат, энергия алмаса алатын макроскопиялық денелер жиынтығын айтады. Егер осындай алмасулар жүйеге енетін денелер арасында ғана болса, ондай жүйе оңашаланған жүйе деп аталады. Егер жүйе сыртқы ортамен осындай алмасулар жасайтын болса, ондай жүйе ашық жүйе деп аталады. Ал жүйе өзін қоршаған ортамен тек қана энергия алмаса алатын болса, онда оны тұйық жүйе деп атайды. Жүйенің күйі макроскопиялық физикалық шамалардың өзгеруін көрсететін параметрлермен беріледі. Маңызды параметрлерге көлем, температура, қысым, сонымен қоса электрлік поляризация, магниттеліну және т. б. жатады. Жүйенің жалпылама күй теңдеуі
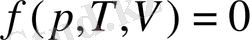
Күй теңдеуіне мысал ретінде идеал газға арналған Менделеев- Клапейрон теңдеуін алуға болады:
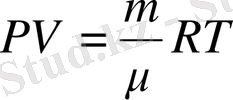
Тепе-теңдік күй немесе термодинамикалық тепе-теңдік күйге дене жеткілікті уақыт өткеннен кейін өздігінен өзі келеді, егер жүйе оңашаланған болса. Тепе-теңдік күйде барлық қайтымсыз процестер тоқталады, жүйе күйін сипаттайтын параметрлер уақытқа байланысты өзгеріссіз қалады. Жүйенің бір күйден екінші күйге өтуі термодинамикалық процесс деп аталады.
Бірақ табиғаттағы барлық процестер қайтымсыз процестер. Энергияның біраз бөлігі жылуға айналатын болса, ондай процесс қайтымсыз, себебі жылу қайтадан энергияға айнала алмайды. Сонымен, кез келген процесс қайтымды болу үшін үйкеліс болмауы керек, Қайтымсыз процестер тек бір ғана бағытта жүреді.
Ішкі энергия жүйеге кіретін барлық бөлшектердің кинетикалық және потенциалық энергияларының қосындысы. Термодинамикада ішкі энергия ұғымының өзі емес, оның өзгерісі басты роль атқарады.
Энтропия энергияның қайтымсыз шашырауының өлшем бірлігі және термодинамикалық жүйе күйі функциясы болып табылады. Процестерді зерттеген кезде оның өзгерісі ерекше қызығушылық тудырады. Энтропия мынаған тең
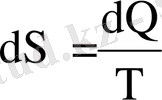
Қайтымды процестер үшін энтропия тұрақты, ал қайтымсыз процестер үшін өседі.
Сонымен,
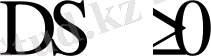
Бұл Клуазиус теңсіздігі. Энтропия жүйедегі ретсіздік өлшемі болып табылады. Оны былайша да өрнектеуге болады:
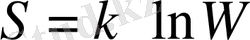
Мұндағы
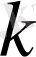 -Больцман тұрақтысы,
-Больцман тұрақтысы,
 -жүйе күйінің термодинамикалық ықтималдылығы. Бұл Больцман формуласы деп аталады. Жүйені макрокүйге түсіру қанша микро күйден өткенін көрсететін санды жүйе жүйенің термодинамикалық ықтималдығы деп атайды.
-жүйе күйінің термодинамикалық ықтималдылығы. Бұл Больцман формуласы деп аталады. Жүйені макрокүйге түсіру қанша микро күйден өткенін көрсететін санды жүйе жүйенің термодинамикалық ықтималдығы деп атайды.
Еркін энергия
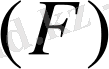 бұл жүйенің ішкі энергиясының бір бөлігі, осының есебінен жүйе қайтымды изотермиялық процесте жұмыс жасауы мүмкін.
бұл жүйенің ішкі энергиясының бір бөлігі, осының есебінен жүйе қайтымды изотермиялық процесте жұмыс жасауы мүмкін.
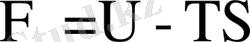
мұндағы
 шамасын кейде байланысқан энергия деп те атайды. Егер жүйе тепе-теңдік күйге жетсе, онда еркін энергия минимал, ал энтропия максимал болады. Термодинамикалық тепе-теңдік күйде барлық макроскопиялық процестер тоқтайды да, жүйедегі денелерге ортақ температура орнайды.
шамасын кейде байланысқан энергия деп те атайды. Егер жүйе тепе-теңдік күйге жетсе, онда еркін энергия минимал, ал энтропия максимал болады. Термодинамикалық тепе-теңдік күйде барлық макроскопиялық процестер тоқтайды да, жүйедегі денелерге ортақ температура орнайды.
Термодинамиканың бірінші заңы энергияның сақталу заңы болып табылады. Жүйеге берілген жылу оның ішкі энергиясын арттыруға және сыртқы күштерге қарсы жұмыс істеуге кетеді:
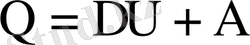
Егер жүйе бернеше күйлерде бола отырып, өзінің алғашқы күйіне қайтып келсе, онда ішкі энергия нолге тең

Сонымен жүйе оған берілген жылу мөлшері есебінен ғана жұмыс жасай алады.
Термодинамиканың бірінші заңы табиғатта болып жатқан процестердің бағыты жөнінде ешқандай мағлұмат бермейді. Термодинамиканың екінші заңы өмірде болатын процестердің бағыты жөнінде мағлұмат береді. Анықтамалары:
«Жылу ешқашан да суық денеден ыстық денеге берілмейді».
«Мәңгі двигательдің екінші түрін жасау мүмкін емес».
Термодинамика заңдарын тірі табиғатқа да қолдануға болады. Организмге келіп түскен тамақтан пайда болатын энергияның мөлшері организм жұмыс істегенде кететін энергияның мөлшеріне тең екені анықталды. Шамамен алғанда берілген энергия (7854 кДж) денеден бөлінген энегияға (7771 кДж) тең екен. Олай болса организм энергияның жаңа көзі болып саналмайды екен. Осыдан келіп, термодинамиканың бірінші бастамасы биологиялық жүйелерге де жарай береді деген қорытындыға келеміз. Жан-жануарлардың организмінен бөлініп шыққан энергия оны қоршаған ортаға таралып кетеді.
Жан-жануар организміне түскен тамақтың күрделі биохимиялық реакцияға түсетіні мәлім. Соның нәтижесінде ол тамақ әсерінен қанша энергия бөлініп шығатынын қалай табуға болады дегне заңда сұрақ туады. Бұл сұраққа 1836 жылы ашылған Гесс заңы жауап береді.
Көптеген сатылардан өтіп келген химиялық реакцияның жылулық эффектісі реакцияның жүріп өткен жолына байланысты болмайды, ол тек қана химиялық жүйенің бастапқы күйдегі энергиясы мен соңғы күйдегі энергиясының айырмасына байланысты болады:

Мұндағы
 - реакцияның бастапқы энергиясы
- реакцияның бастапқы энергиясы
 -реакцияның соңғы энергиясы
-реакцияның соңғы энергиясы
Ол үшін тамақты өртейді. Ол құралды калориметриялық бомба деп атайды. Сонда бөлініп шыққан жылуды өлшеп алады. Ал осы тамақты адам жесе, онда тамақ организмде биохимиялық реакцияға түсіп, жылу бөліп шығарады. Гесс заңы бойынша осы екі жылу біріне-бірі тең болуға тиіс.
Тірі организмдердегі энергияның түрленулері
«Термодинамикалық тепе-теңдік күй»
1 Термодинамикалық стационар күй.
Термодинамика заңдарын биологиялық жүйелерге қолданғанда тірі организмнің ерекшеліктеріне аса көңіл бөлу керек: 1) заттар мен энергия ағынына биологиялық жүйелер ашық; 2) Тірі жүйелердегі процесстер қайтымсыз; 3) тірі жүйелер тепе-теңдіктен алыс; 4) биологиялық жүйелер гетерофазалы, құрылымдық және жеке фазалары аздаған молекулалар санынан тұруы мүмкін. Биологиялық жүйелер қасиеттерін нақты түрде сипаттау үшін қайтымсыз процестер термодинамикасы теориясы қолданылады. Оның негізіг салушылар Л. Онгазер мен И. Пригожин. Процестің уақытқа тәуелділігіне мысал ретінде:
 иондарының концентрациясы клетканың ішіне қарағанда сыртында көп. Бірақ концентрациясы градиенті мен потенциалодар айырмасының болуы
иондарының концентрациясы клетканың ішіне қарағанда сыртында көп. Бірақ концентрациясы градиенті мен потенциалодар айырмасының болуы
 индарының ішке еніп кетуіне әкеледі, сондықтан концентрациясы тұрақты болып қалады.
индарының ішке еніп кетуіне әкеледі, сондықтан концентрациясы тұрақты болып қалады.
Стационарлық күй сипаттамалары:
Жүйеге енген зат мен одан бөлініп шыққан заттың тұрақтылығы;
Еркін энергия шығынының тұрақтылығы, олар жүйедегі заттар концентрациясын тұрақты етіп ұстап тұрады.
Стационар күйдегі термодинамикалық параметрлердің тұрақтылығы.
Ашық жүйе стационар күйде зат мен электр зарядтары ағыны есебінен ғана өмір сүре алады.
Сызықты тепе-тең емес термодинамика негізін қалаған Л. Онзагер мен И. Пригожин болды. Ол тепе-тең күйге жақын процестерді қарастырады, процестерден пайда болған жылдамдықтар мен күштердің арасында сызықтық байланыстар болған кездегі.
Биологиялық жүйелер градиент санының артықтығымен сипатталады (осмостық, электрлік, концентрлік және т. б.
Қандай да бір термодинамикалық параметрдің градинті ара қашықтыққа байланысты өзгереді.
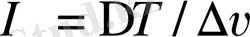
 - үлкен параметрден кіші параметрге дейінгі бағыт.
- үлкен параметрден кіші параметрге дейінгі бағыт.
Биологиялық жүйе оның градиенті болса, онда оның жұмыс істей алу мүмкіндігі бар. Градиентті энергия қоймасы деп айтуға болады.
Еркін энергия
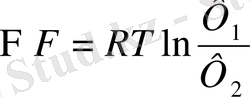
 - термодинамикалық параметрлердің 1-ші және 2-ші нүктелердегі мәні.
- термодинамикалық параметрлердің 1-ші және 2-ші нүктелердегі мәні.
Жүйенің жұмыс істеуі осы еркін энергияны жұмсауына байланысты. Егер жұмыс істелетін болса, онда градиент кемиді, себебі оның энергиясы есебінен жұмыс жасалады, бірақ параллель оған қарама-қарсы бағытта екінші градиент пайда болады. Қайтымсыз процестерде екінші градиенттің шамасы бірінші градиентке қарағанда аз болады.
2 Биологиялық процестердің п. ә. к.
ПӘК- істелген жұмыстың осы жұмысты істеуге кеткен еркін энергия өзгерісіне қатынасы:
ПӘК
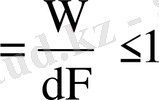
ПӘК абсолю бірлікпен немесе процентпен берілуі мүмкін. Термодинамиканың екінші бастамасына сәйкес қайтымды процестердің ПӘК бірге тең, ал қайтымсыз процестердікі бірден кіші. Ал нақты биологиялық процестердің ПӘК-і де бірден кіші. Нақты биологиялық процестердің ПӘК жуық шамамен мынаған тең:
Гликолиз - 36%
Ф/с -75%
Қышқылдандыратын фосфорлану - 55%
Бұлшық еттің жиырылуы - 40%
Стационар күй дегеніміз термодинамикалық жүйенің параметрлері уақытқа байланысты өзгермейді, бірақ зат алмасу мен энергия алмасу жүріп жататын күй. Ашық жүйелер термодинамикалық тепе-тең күйлерге өтуі мүмкін.
Стационар күй:
- Қоршаған ортамен тұрақты түрде энергия алмасу
- Күйді сақтап тұру үшін еркін энергия тұрақты түрде жұмсалып отырады
- Термодинамикалық потенциал тұрақты.
- энтропия тұрақты, бірақ максимал емес
- градиенттер болмайды.
Термодинамикалық тепе-теңдік
- қоршаған ортамен зат және жылу алмасу болмайды
- осы күйді ұстап тұру үшін еркін энергия жұмсалмайды
- жүйенің жұмыс істей алу қабілеті мен термодинамикалық потенциалы нольге тең.
- энтропия максималь
- жүйеде градиенттер болмайды.
Пригожин теоремасы
Кез келген ашық термодинамикалқы жүйеде тұрақты түрде энтропия пайда болады, оның ішінде биожүйелерде де.
Пригожин анықтамасы:
Сыртқы параметрлер белгілі болған стационар күйлерде ашық термодинамикалық жүйелерде энтропия өзінің минимал мәніне ұмтылады. Энтропия - еркін энергияны жұмсау өлшемі, сондықтан кез келген ашық термодинамикалық жүйе стационар күйде еркін энергияны минимал жұмсайды.
Ле-Шателье-Браун принципі
Егер орнықты тепе-теңдік күйде тұрған жүйеге сырттан қандай да бір параметрін (температура, қысым, концентрация) өзгерте отырып әсер етсек, онда жүйеде сыртқы әсерлерді компенсациялайтын процестер арта бастайды.
Биоэлектрлік потенциалдар.
- Биоэлектрлік потенциалдардың сипаттамалары және пайда болуының негізгі себептері.
Биоэлектрлік потенциалдар деп жануарлардың, адамдардың тканінде, клеткаларында пайда болатын потенциалдар айырмасын айтамыз. Тірі организмдегі потенциалдар айырмасының қалай пайда болатынын түсіну үшін ерітінділердегі потенциалдардың қалай пайда болатынын қарастыра кетейік.
Ерітінділерде пайда болатын потенциалдар электронды және ионды болып екіге бөлінеді. Айталық, ерітіндіге мыс және және мырыш салынсын. Мыс еріген кезде ьос электрондар бөліп шығарып, өзі оң зарядталса, мырыш ол электрондарды қабылдайды да теріс зарядталады. Сөйтіп ерітіндіде потенциалдар айырымы пайда болады. Ерітінділердегі иондардың әсерінен пайда болатын потенциалдарды ион типті потенциалдар деп атайды. Ион типті потенциалдар диффузиялық, мембраналық және фазааралық болып үшке бөлінеді.
Диффузиялық потенциалдар айырмасы Гендерсон формуласымен анықталады:
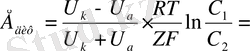
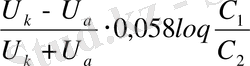
Мұндағы
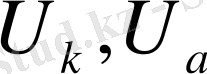 - катион мен анион қозғалғыштығы,
- катион мен анион қозғалғыштығы,
 -Кельвин шкаласындағы температура,
-Кельвин шкаласындағы температура,
 - универсаль газ тұрақтысы,
- универсаль газ тұрақтысы,
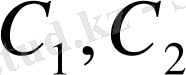 - сұйықтар концентрациясы,
- сұйықтар концентрациясы,
 иондар валенттілігі.
иондар валенттілігі.
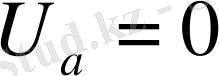 болса, онда Гендерсон теңдеуі мына түрге келеді:
болса, онда Гендерсон теңдеуі мына түрге келеді:
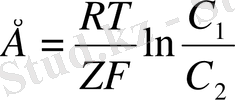
Бұл Нернст теңдеуі деп аталады.
Клеткалардың цитоплазмасында және сыртқы ортада негізінен калий
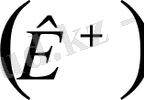 , натрий
, натрий
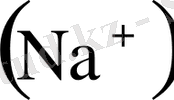 , хлор
, хлор
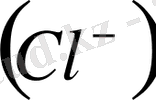 иондар мен аминқышқыл аниондары
иондар мен аминқышқыл аниондары
 жүреді. Клетканың ішінде калий, хлор және аминқышқыл аниондары, ал клетканың сыртында калий
жүреді. Клетканың ішінде калий, хлор және аминқышқыл аниондары, ал клетканың сыртында калий
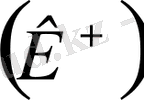 , және хлор
, және хлор
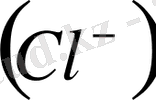 иондары бар дейік. Калий және хлор иондары мембрананың ішкі және сыртқы жағына еркін өтіп жүретін болса, аминқышқыл анионы клетка ішінде қалып қояды. Мембрананың ішкі және сыртқы жағындағы иондардың концентрациясының көбейтіндісі біріне-бірі тең болады да, оны Доннан тепе-теңдігі деп атайды, яғни:
иондары бар дейік. Калий және хлор иондары мембрананың ішкі және сыртқы жағына еркін өтіп жүретін болса, аминқышқыл анионы клетка ішінде қалып қояды. Мембрананың ішкі және сыртқы жағындағы иондардың концентрациясының көбейтіндісі біріне-бірі тең болады да, оны Доннан тепе-теңдігі деп атайды, яғни:
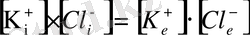 немесе
немесе
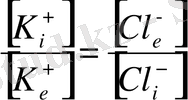
мұндағы
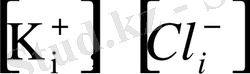 - мембрананың ішіндегі калий және хлор иондары, ал
- мембрананың ішіндегі калий және хлор иондары, ал
 - мембрананың сыртындағы калий және хлор иондары. Сонда Нернст теңдеуі былайша өрнектеледі:
- мембрананың сыртындағы калий және хлор иондары. Сонда Нернст теңдеуі былайша өрнектеледі:
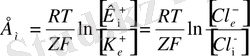
Фазааралық потенциалдар араласпайтын екі сұйықтың шекарасында потенциалдар пайда болады. Клетканың цитоплазмасын көп фазалы микрогетерогенді жүйе деп қарастыратын болсақ, онда фазалар шекарасында потенциалдар айырмасы пайда болады. Оны фазааралық потенциалдар деп атайды.
Электродтық потенциалдлар шамасы мына формуламен анықталады:
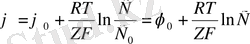
мұндағы
 -металдың электродтық потенциалы,
-металдың электродтық потенциалы,
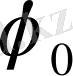 - қалыпты электродтық потенциалы,
- қалыпты электродтық потенциалы,
 - ерітінді концентрациясы,
- ерітінді концентрациясы,
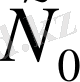 - металдағы ионның концентрациясы, ол
- металдағы ионның концентрациясы, ол
 , бұл Нернст теңдеуі деп аталады. Қалыпты электродтық потенциалдың шамасы
, бұл Нернст теңдеуі деп аталады. Қалыпты электродтық потенциалдың шамасы
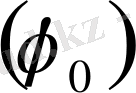 кестеден алынады. Кейбір металдардың қалыпты электродтық потенциалы мына кестеде берілген.
кестеден алынады. Кейбір металдардың қалыпты электродтық потенциалы мына кестеде берілген.






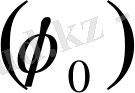

2. Тыныштық потенциалы .
Жануарлар мен адамдардың клеткаларында, тканінде биоэлектрлік потенциалдар пайда болады дедік. Бұл биоэлектрлік потенциалдар тыныштық потенциалы, әсер потенциалы, постинаптикалық және генераторлық болып бөлінеді. Тыныштық потенциалын мембраналық тыныштық потенциалы деп те атайды. Тірі клетканың тыныштық күйінде тұрғанда оның (клетканың) ішіндегі зат пен сыртындағы ерітіндінің арасында потенциалдар айырмасы пайда болады. Оны тыныштық потенциалы деп атайды. Тыныштық потенциалының шамасы 50-100 В тең. Бұл потенциал мембрананың бетіне жиналады. Мембрананың ішкі беті сыртқы бетіне қарағанда теріс зарядталады. Тыныштық потенциалының пайда болуы мембрана арқылы калий иондарының
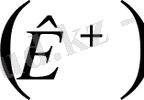 өзіне байланысты болады. Протоплазмадағы калий
өзіне байланысты болады. Протоплазмадағы калий
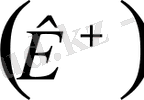 концентрациясы клетка сыртындағы сұйық ішіндегі концентрацияға қарағанда елу есе көп болады да, мембрана арқылы сыртқа оң зарядталған калий иондары шығады. Сөйтіп мембрананың ішкі беті теріс зарядталады. Тыныштық потенциалы Нернст формуласымен анықталады.
концентрациясы клетка сыртындағы сұйық ішіндегі концентрацияға қарағанда елу есе көп болады да, мембрана арқылы сыртқа оң зарядталған калий иондары шығады. Сөйтіп мембрананың ішкі беті теріс зарядталады. Тыныштық потенциалы Нернст формуласымен анықталады.
Егер клетканың белгілі бір әдіспен тітіркендірсек, ол қозады. Бұл кезде клетканың физикалық, химияық, функциялық өзгеріске ұшырайды. Қозған кезде клетка мембранасының электрлік күйі өзгеріске түседі. Клетканың қозған жері оның қозбаған жеріне қарағанда теріс зарядталады. Олай болса, қозған жердегі иондар қозғалысқа түсіп басқа жерге орын ауыстырады деген сөз. Клеткаға әсер ететін тітіркендіргіштер алдымен тыныштық потенциалының төмендеуіне әкеліп соқтырады. Тыныштық потенциалы төменгі шегіне жеткеннен кейін әсер потенциалының активті (белсенді) кезеңі басталады. Әсер потенциалы нерв (жүйке) тамырларымен немесе бұлшық ет тарамдарымен өте үлкен жылдамдықпен таралады. Әсер ету ұзақтығы 2 миллисекундқа тең болады.
3. Тірі жүйелерде электрлік потенциалдардың айырмашылықтарын тіркеп алу әдістемелері .
Клетканың ішкі және сыртқы беттеріндегі потенциалдарды өлшеу үшін өте нәзік әдіс қажет. Потенциалдарды өлшеу үшін микроэлектродтық әдіс деген қолданылады. Шыныдан жасалған диаметрі 0, 5-1 мкм (микрометр) (
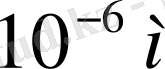 ) пипетканың іші хлорлы калий
) пипетканың іші хлорлы калий
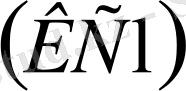 ерітіндісімен толтырылады. Клетканың ішінде калий иондары болатын ерітінді алынады. Екінші жағынан бұл ерітінді электродтың өз потенциалын өте аз мөлшерде туғызады. Пипетканың ұшының диаметрі 0, 5 мкм болса, онда оның өз кедергісі 10-30 мОм болады. Микроэлектродтың өз потенциалы болады. Алайды бұл потенциал тірі тканьнің потенциалдар айырмасын дәл өлшеуге онша кедергі жасамайды. Микроэлектродты жануарлар тканіне енгізу микроманипулятордың көмегімен іске асады. Бұл микроманипулятор үш шприцтен тұрады. Оның біреуі микроэлектродқа жалғанады. Екінші электрод клетканың сыртқы бетіне жапсырылады. Сөйтіп дененің іші мен сыртындағы потенциалдар айырымы өлшенеді. Организмдегі әртүрлі клеткалар, тканьдер қызмет атқарады. Олардың электр активтілігі электр өрісінің пайда болуына мүмкіндік жасайды. Потенциалдар айырмасының уақыт бойынша өзгеруін электрограмма дейді. Бұл электрограмманы дененің қай органы шығарып тұруына байланысты әр түрлі атайды. Жүректін биопотенциалдарының уақыт бойынша өзгеруін электрокардиограмма (ЭКГ), бұлшық ет потенциалының уақыт бойынша өзгеруін электромиграмма (ЭМГ), ал ми қабының биопотенциалының уақыт бойынша өзгеруін электроэнцефалограмма (ЭЭГ) делінеді.
ерітіндісімен толтырылады. Клетканың ішінде калий иондары болатын ерітінді алынады. Екінші жағынан бұл ерітінді электродтың өз потенциалын өте аз мөлшерде туғызады. Пипетканың ұшының диаметрі 0, 5 мкм болса, онда оның өз кедергісі 10-30 мОм болады. Микроэлектродтың өз потенциалы болады. Алайды бұл потенциал тірі тканьнің потенциалдар айырмасын дәл өлшеуге онша кедергі жасамайды. Микроэлектродты жануарлар тканіне енгізу микроманипулятордың көмегімен іске асады. Бұл микроманипулятор үш шприцтен тұрады. Оның біреуі микроэлектродқа жалғанады. Екінші электрод клетканың сыртқы бетіне жапсырылады. Сөйтіп дененің іші мен сыртындағы потенциалдар айырымы өлшенеді. Организмдегі әртүрлі клеткалар, тканьдер қызмет атқарады. Олардың электр активтілігі электр өрісінің пайда болуына мүмкіндік жасайды. Потенциалдар айырмасының уақыт бойынша өзгеруін электрограмма дейді. Бұл электрограмманы дененің қай органы шығарып тұруына байланысты әр түрлі атайды. Жүректін биопотенциалдарының уақыт бойынша өзгеруін электрокардиограмма (ЭКГ), бұлшық ет потенциалының уақыт бойынша өзгеруін электромиграмма (ЭМГ), ал ми қабының биопотенциалының уақыт бойынша өзгеруін электроэнцефалограмма (ЭЭГ) делінеді.
Жүрек - дипольдің электр моменті уақыт бойынша өзгеріп отырады, олай болса, тарамдарда кернеудің уақыт бойынша байланысы өрнектеледі. Электрокардиограмманы жазып алып, жүректің жұмысын тексеруге болады.
Биологиялық жүйелерде тұрақты және айнымалы токтардың өту заңдылықтары
1. Биологиядағы Ом заңы. Поляризация құбылысы.
Қатты дене, сұйық және газ меншікті электр өткізгіштігімен сипатталады. Дененің электр өткізгіштігі деп уақыт бойынша өзгермейтін электр өрісі әсерінен заттың электр тогын өткізу қабілетін айтамыз. Денелерде ток тасымалдаушылар болады. Мысалы, металдарда, жартылай өткізгіштерде ток тасымалдаушылар электрондар болса, электролиттерде иондар, ал плазмаларда электрондар мен иондар болып саналады. Меншікті электрөткізгіштігіне қарап денелерді өткізгіштер
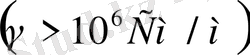 , жартылай өткізгіштер
, жартылай өткізгіштер
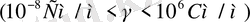 және диэлектриктер
және диэлектриктер
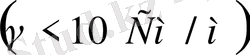 деп үшке бөлінеді. Иондық өткізгіштердің меншікті электр өткізгіштігі
деп үшке бөлінеді. Иондық өткізгіштердің меншікті электр өткізгіштігі
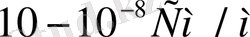 аралығында жатады. Ал биологиялық объектілер өткізгіштің де, диэлектриктің де қасиетіне ие бола алады. Клеткалар мен ұлпаьдердегі бос иондар олардың электр өткізгіштігін қамтамасыз етеді. Тірі организмдегі электр тогын электрон, иондар алып жүреді. Ом заңы тірі организмде сол күйінде орындалмайды. Өткізгіш арқылы өтетін токтың шамасы кернеуге тура пропорционал екені белгілі:
аралығында жатады. Ал биологиялық объектілер өткізгіштің де, диэлектриктің де қасиетіне ие бола алады. Клеткалар мен ұлпаьдердегі бос иондар олардың электр өткізгіштігін қамтамасыз етеді. Тірі организмдегі электр тогын электрон, иондар алып жүреді. Ом заңы тірі организмде сол күйінде орындалмайды. Өткізгіш арқылы өтетін токтың шамасы кернеуге тура пропорционал екені белгілі:
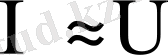 немесе
немесе
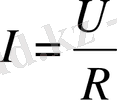 . Бұл Ом заңы. Тірі организм арқылы ток өтіп жатсын. Оған берілетін кернеу өзгермесе де, одан өтетін ток өзгереді. Дәлірек айтқанда ток күші кемиді. Ток күшінің кемуі -ұлпада болып жатқан поляризацияға байланысты болады. Атап айтқанда, ұлпалардың сиымдылық, диэлектрлік қасиеттерінен ток күші азаяды. Биологиялық объектілер үшін Ом заңы былайша өрнектеледі:
. Бұл Ом заңы. Тірі организм арқылы ток өтіп жатсын. Оған берілетін кернеу өзгермесе де, одан өтетін ток өзгереді. Дәлірек айтқанда ток күші кемиді. Ток күшінің кемуі -ұлпада болып жатқан поляризацияға байланысты болады. Атап айтқанда, ұлпалардың сиымдылық, диэлектрлік қасиеттерінен ток күші азаяды. Биологиялық объектілер үшін Ом заңы былайша өрнектеледі:
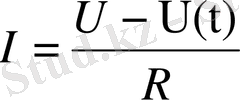
мұндағы
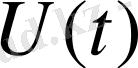 поляризация қозғаушы күші.
поляризация қозғаушы күші.
Диэлектриктер кез келген зат сияқты атомдар мен молекулалардан тұрады. Оң заряд атом ядросында, теріс заряд атомдар мен молекулалардың электрондық қабықшаларында жинақталған. Жалпы алғанда, оң және теріс зарядтар өзара тең, сондықтан атом (молекула) электрлік нейтрал. Молекуланы электрлік диполь ретінде қарастыруға болады. Диэлектрикті сыртқы электр өрісіне орналастырсақ, ол поляризацияланады да, оның дипольдік моменті мынаған тең болады:
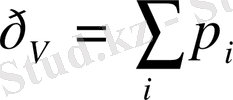 , мұндағы
, мұндағы
 -бір молекуланың дипольдік моменті. Диэлектрик поляризациясы дегеніміз сыртқы өріс әсерінен диэлектрик дипольдарының орналасуын, сөйтіп электр қозғаушы күшінің пайда болуын айтамыз.
-бір молекуланың дипольдік моменті. Диэлектрик поляризациясы дегеніміз сыртқы өріс әсерінен диэлектрик дипольдарының орналасуын, сөйтіп электр қозғаушы күшінің пайда болуын айтамыз.
Әртүрлі зарядталған екі шексіз параллель жазықтықтар туғызған біртекті сыртқы электр өрісі арасына диэлектрик қояйық (1-сурет) .
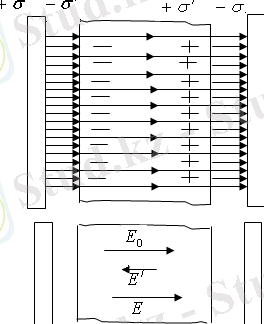
(1-сурет)
Өріс әсерінен диэлектрик поляризацияланады, зарядтардың ығысуы пайда болады: оң зарядтар өріс бойымен, теріс зарядтар өріске қарсы. Осының нәтижесінде диэлектриктің оң қырында көлемдік тығыздығы
 болатын оң зарядтар, ал сол қырында көлемдік тығыздығы
болатын оң зарядтар, ал сол қырында көлемдік тығыздығы
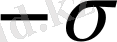 болатын теріс зарядтар артық болады. Осы поляризациядан пайда болған компенсирленбенген зарядтар байланысқан зарядтар деп аталады. Олардың беттік тығыздығы
болатын теріс зарядтар артық болады. Осы поляризациядан пайда болған компенсирленбенген зарядтар байланысқан зарядтар деп аталады. Олардың беттік тығыздығы
 жазықтықтың еркін зарядтарының тығыздығы
жазықтықтың еркін зарядтарының тығыздығы
 -дан кем болады. Өрістің кернеулік сызықтарының бір бөлігі диэлектриктен өтіп кетеді, ал қалған бір бөлігі байланысқан зарядтарда үзіліп қалады. Сондықтан, диэлектрик поляризациясы өрісті алғашқы сыртқы өріспен салыстырғанда кемітеді. Диэлектриктен тыс жерде
-дан кем болады. Өрістің кернеулік сызықтарының бір бөлігі диэлектриктен өтіп кетеді, ал қалған бір бөлігі байланысқан зарядтарда үзіліп қалады. Сондықтан, диэлектрик поляризациясы өрісті алғашқы сыртқы өріспен салыстырғанда кемітеді. Диэлектриктен тыс жерде
 .
.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz