Жасушаның құрылысы мен тіршілік циклінің негіздері: зат алмасу және қалқанша безінің гормондық реттелуі

Жоспар
І. Кіріспе
ІІ. Негізгі бөлім
Жасушаның тіршілік циклі, құрылысы
Жануарлар мен өсімдіктердің жасушасы
ІII. Пайдаланылған әдебиеттер тізімі
Пайдаланылған әдебиеттер тізімі:
1. Нұрышев М., «Гистология және эмбринология негіздері» Алматы, 1998ж.
2. Нұрышев М. Х., Нұрышева А. М. «Цитология» Алматы, 1999ж.
3. Антипчук Ю. П. Гистология с основами эмбриологии. 1993.
4. Манушюва Н. А. Гистология с основами эмбриологии. М., 1983.
5. Рақышев А. Организмнің нәзік құрылымы. Алматы, 1994.
6. Сапаров К. Жалпы цитология негіздері. Алматы, 1994.
7. Токин Б. П. Общая эмбриология. М., 1987.
8. Хэм А., Кормак Д. Гистология. т. I-III, 1993.
9. Ченцов Ю. Общая цитология. М., 1994.
Кіріспе
Жасушаның атомдық және молекулалық құрамы. Табиғатта кезде-сетін 105 химиялық элементтің 80-ге жуығы тірі ағза жасу-шаларының құрамында болады. Олар жасушадағы мөлшеріне карай үш топқа бөлінеді:
1. Органикалық қосылыстардың құрамына кіретін негізгі элементтер. Олардың жасуша күрамындағы үлесі 99%;
2. Ион түріңде кездесетін элементтер. Олардың жасушадағы жалпы мөлшері 1% шамасында;
3. Жасушада өте аз мөлшерде кездесетін (0, 01%-тен кем) микро-элементтер.
Жасушада кездесетін элементтер 1-кестеде берілген. Осы кестеде берілген элементтердің құрамы барлық жануарлар жасушасында бірдей болады, ал өсімдіктер мен ұсақ ағза жасушаларында басқаша. Тірі ағзадағы химиялық элементтердің зат айналым әрекетіңдегі маңызы соншалық, егер микроэлементтер жетіспесе, ағза ауруға ұшырайды. Мәселен, жасушадағы иодтың мөлшері 0, 01% болуы қажет. Егер тағамның құрамыңда иод жетіспесе, бала бойының өсуі тежеледі. Егер тағамның құрамыңда мыстың мөлшері жетіспесе, мал жаппай ауруға ұшырайды. Сонымен қатар, химиялық элементтердің тірі ағзалардағы үлесі жер бетіндегі таралуынан мүлдем өзгеше. Тірі ағза жасушасыңда (клеткасында) кездесетін сутегі, оттегі, көміртегі және азоттың үлесі 99%-ке дейін жетеді. Осылардың үшеуінің (сутегі, көміртегі, азот) жер бетіндегі жалпы үлесі 1%-тен кем болады.
Жасушаның тіршілік циклі, құрылысы.
Тіршіліктің мәнін анықтау - биологияның негізгі мақсаттарының бірі.
Тіршілік анықтамасы оте көп, бірак солардың ішінде кең таралғаны Ф. Энгельстің берген анықтамасы болып саналады. Ф. Энгельс "Тіршілік - ақуыз денелердің омір сүру формасы, оның негізгі мәні сыртқы ортамен зат алмасуы" деп керсеткен болатын.
Қазіргі кездегі көзқарас бойынша тіршіліктің негізін 2 ірі биополимерлер-ақуыз және нуклеин қышқылдары құрайды. Сондықтан да тіршіліктің барлық құбылыстары осы екі биоголимерлердің кешеңді қасиеттеріне байланысты.
Тірі ағзалар өлі дүниеде кездесетін химиялық элементтерден тұрады, бірақ сол химиялық заттардын тірі ағзалардағы күйі, атқаратын қызметі, активтігі өлі дүниедегіден өзгеше болады.
Тірі ағзалар денесінде заттардың тек физико-химияық өзгерістері мен күрделі физиологиялық құбылыстары жүріп коймай, сол сияқты онда сапалық биологиялық заңдылықтар байқалады (ыдырау-синтезделу; заттар, энергия, ақпараттар ағыны т. б. ) .
Тіршіліктің түпкілікті қасиеттеріне мыналар жатады:
өздігінен жаңару - ол заттар мен энергия ағыңдарына байланысты;
өздігінен кобею - бұл бірін-бірі алмастырып отыратын ұрпақтар жалғасын қамтамасыз ететін процесс және ол ақпараттар ағынына байланысты болады;
өздігінен реттелу - ол заттардың, энергия және ақпараттар ағындарына байланысты.
Тіршіліктің жоғарыда келтірілген түпкілікті қасиеттері оның негізгі белгілерін қалыптастырады, олар: заттардың және энергияның алмасуы, тітіркену, гомеостаз, көбею, тұқым куалау мен өзгергіштік, жеке және филогенетикалық даму, тіршіліктін дискреттілігі және біртұтастығы.
1) Заттардың және энергияның алмасуы. Заттардың алмасуы
тіршіліктің ең негізгі белгілерінін бірі болып саналады. Ол кейде қара-пайым күйде өлі дүниеде де кездеседі. Бірақ ол тірі ағзалардан өзгеше жүреді, мысалы: көмір жанғанда 0
2
пен С0
2
алмасады, темірдің бетін тат басқанда да 0
2
алмасуы байқалады. Өлі дүниеде бұл процестер негізінде заттар ыдырап жойылып жоқ болады, ал тірі ағзалардын тіршілік етуі үшін міндетті түрде зат алмасу болуы кажет. Зат аламасу процесіңде тірі ағзаларда ыдыраған заттар жойылмай қайтадан калпына келіп отырады,
яғни өздігінен жанару және ездігінен көбею байқалады.
Тірі ағзалар ашық биологиялық жүйе болып табылады. Ол арқылы үзіліссіз заттар, энергия және ақпараттар ағыны өтіп отырады.
XVIII ғасырда Лавуазье жану процесімен тірі клеткадағы тотығу процестерінің арасында ұқсастық бар екендігін айтты. 1898 жылы Бах А. Ң. асқын тотық теориясын ұсынды, яғни оттегі тотығушы заттардьщ энергріясының арқасында белсендіріледі, сөйтіп пероксидтер түзіледі.
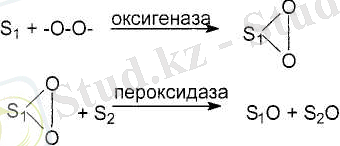
Дегенмен, бұл мысал кейінгі зерттеулерге қарағанда биологиялық тотығудың микросомаларда журетін бір ғана мысалы бола алады.
тотығу процесінің соңғы өнімі су болады деп есептеп, цитохромоксидаза ферментін ашты.
1912 жылы Палладин биототығудың жаңа түсініктерін еңгізді, яғни биототығу аэробты және анаэробты кезеңдерден тұрады, анаэробтық тотығуда субстрат сутектен айырылады.
1933 жылы Кейлин электрондары тасымалдаушы цитохромдарды ашты. Оксигеназалық және дегидрогеназалық теориялар клеткадағы биологиялық тотығудың немесе үлпалық тыныс алудың негізі болып табылады. Онан әрі қарай, отандық және шетелдік ғалымдардың жүмыстарының нәтижесінде биологиялық тотығу - субстраттан көптеген ферменттік жүйелердің көмегімен электрондардың, протондардың оттегіне жеткізілуінің арқасында су және энергия түзілетін көп сатылы тотығу-тотықсыздану процестері екендігі, ал бүл процестер кезіндегі бөлінген энергия АТФ фосфатының байланысқан энергиясына айналатындығы деп дәлелденеді.
Сутегінің оттегімен тотығуынан 1 моль су түзілген кезде 230 кДж энергия бөлінеді де, осы энергияның

біразы АТФ-тың түзілуіне жүмсалады
1. Субстраттық фосфорлану - АТФ-тың синтезі макроэргтік қосылыстың энергиясының
арқасында түзіледі, оттегінің қажеті жоқ.
2. Тотығып фосфорлану - АТФ синтезінің негізгі жолы. Барлық АТФ-тың 90%-не дейін осы жолмен синтезделеді. Протондармен электрондардың НАДН 2 пен ФАДН 2 -ден оттегіне дейін электрондарды хасьшалдау тізбегі арқылы жеткізу кезінде АТФ синтезінің катарласып, ідесіп жүруі. Электрондар мен протондардың доноры - органикалық субстраттар (SH 2 ), ал акцепторлары молекулалық оттегі.
2) Тітіркену - тіршіліктің негізгі белгілерінің бірі. Ол сыртқы ортадан берілетін кез келген ақпаратка тірі ағзалардың кері жауап реакциясы болыггсаналады. Сыртқы ортадан қажетті ақпарат алу арқылы биологиялық жүйелердің (жасуша, ұлпа, мүше, ағза т. б. ) өздігінен реттелуі қамтамассыз етіледі.
Өздігінен реттелу негізінде ағзалардын күрылым тұрақтылығы гомеостаз біркалыпты сақталынады.
Көбею - ағзалардың негізгі белгілерінің бірі, онсыз тіршілік болмайды, ұрпақтар жалғаспайды.
Тұқым қуалаушылык пен өзгергіштік - тірі ағзалардың басты белгілері болып саналады.
Даму - тіршіліктің ең негізгі белгісі, Ол жеке даму (онто-генез) және тарихи даму (филогенез) болып бөлінеді.
Дискреттік және біртұтастық.
Тіршілік бір жағынан біртұтас, ал екінші жағынан дискретті (бөлшектенген) болып келеді. Тірі дүние біртұтас - бір ағзаның тіршілігі екінші ағза тіршілігімен тығыз байланысты, мысалы: өсімдіктер, өсімдікқоректі жануарлар - жыртқыштар т. с. с. Сондай-ақ, тіршілік жеке-жеке, бір-бірінен дербес бөлшектерден (элементтерден) тұрады, мысалы: өсімдіктер, жануарлар дүниесі түрлерден; түрлер популяциялардан, жеке ағзалардан; ағза-мүшелерден, ұлпалардан, жасушалардан тұрады.
Тіршіліктің дискреттілігі туралы ұғым оны бірнеше құрылым деңгейлерге бөлуге мүмкіндік береді. Қазіргі кезде тіршіліктің 7 ірі құрылым деңгейлері белгілі:
тіршіліктің молекулалық құрылым деңгейі - ДНҚ, РНҚ, ақуыздар (барлық тірі ағзаларда біркелкі болып келеді) ;
тіршіліктің жасушалық құрылым деңгейі (барлык тірі ағзаларда біркелкі болып келеді) ;
тіршіліктің үлпалық қүрылым деңгейі жануарларда - 5, өсімдіктерде 6 түрлі ұлпалар кездеседі;
тіршіліктің ағзалық (онтогенездік) құрылым деңгейі көп түрлі болып келеді;
тіршіліктің популяциялық - түрлік құрылым деңгейі көп түрлі болып келеді; жануарлардың 1, 5 миллионнан астам, есімдіктердің 500 мыңнан астам түрлері белгілі;
тіршіліктің биогеоценоздық құрылым деңгейі - биоценоз дегеніміз бір-бірімен және сыртқы ортамен үнемі қарым-қатынаста болатын тірі ағзалардың тұрақты, тарихи қалыптасқан кауымдастығы;
тіршіліктің биосфералық құрылым деңгейі.
Жасушаның ашылуы оптикалық құрал микроскоптың құрасты-рылуымен тығыз байланысты. Оптикалық әйнектер туралы деректер өте ерте кезден-ақ қалыптаса бастаған. XV ғасырдың басында оптикалық әйнектерді адамдар көзәйнек ретінде пайдаланған. Екі линзадан тұратын және ұсақ денелерді үлкейтіп көрсететін ең қарапайым оптикалық құралды XVI ғасырдың аяғыңда ағайынды Янсендер құрастырған, бірақ ол арқылы жасушаны көруге мүмкіндік болмаған.
Жасушаны көруге мүмкіндік берген алғашқы микроскопты 1665 жылы физик Р. Гук құрастырып, сол микроскоп арқылы жасушаны ашқан. 1671 жылы М. Мальпиги, Н. Грю жасушаларды зерттеп, олардың ең негізгі құрамдық белігі - қабықшасы деп болжамдаған.
XVIII-XIX ғасырларда микроскоптың күрделенуі, микроскопиялык зерттеулер әдісінің жетілуі нәтижесінде бірте-бірте жасушаның тірі заттары ашылған. 1831 жылы ағылшын ғалымы Р. Броун жасушаның түйіршіктенген құрылымын ашып, оны ядро деп атаған; 1841 жылы чех ғалымы Ян Пуркинье жасушаның қоймалжың заты - тірі затын ашып, оны протоплазма деп атаған. Осылайша XIX ғасырда ғалымдардың жасуша туралы пікірі бірте-бірте езгеріп, оның негізгі заты қабықшасы емес, тірі заты - деген ұғым қалыптасқан.
1838-1839 жылдары неміс ғалымдары Т. Шванн және М. Шлейден еңбектерінде өсімдіктер мен жануарлар жасуша-ларының 200 жылға жуык созылған зерттеулері қорытындыланып, жасуша теориясы қалыптасқан. Сол сияқты жасуша теориясының әрі қарай дамуына неміс дәрігері Р. Вирхов та (1858) өз үлесін қосқан.
Қазіргі кезде жасуша теориясының мынадай негізгі қағидалары белгілі:
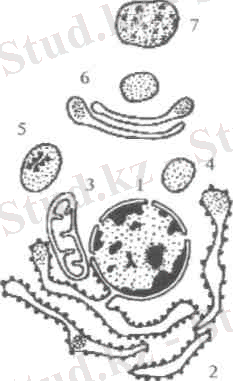
2-сурет. Жасушанын мембраналық жүйесі
1-ядро, 2-кедір-бұдыр эвдоплазмалық тор,
3-цитоплазмалык көпіршік, 4-Гольджи кешені,
5-гранула, 6-митоховдрия, 7-лизосома
эндоплазмалық тор. Гранулалы (кедір-бұдыр) эндоплазмалық тордың мембранасына рибосомалар бекінген, ал тегіс эндоплазмалық торда рибосомалар болмайды. Цитоплазмада, әдетте гранулалы тор агранулалы торға қарағанда әлдеқайда жақсы жетілген. Агранулалы тор кейбір ерекше қызмет атқаратын, яғни майлы заттарды көп синтездейтін жасушаларда ғана жақсы жетілген болады. Гранулалы тордың қызметі - ақуыз синтездеу, жасуша мембраналарын пайда ету орта-лығы болып саналады. Сол сияқты, ол вакуоля, лизосома, микроденешік-терді де пайда ете алады. Эндоплазма-лық арналар арқылы макромолекула-лар, иондар тасымалданады. Аграну-лалы тор липидтерді және кемірсулар-ды синтездеуге қатынасады.
Жасуша орталығы - жануарлар жасушаларына тән органелла. Ол 2 центриолядан тұрады. Әрбір центриоля диаметрі 150 нм, ұзындығы 300-500 нм болып келетін қуыс цилиндр. Оның қабырғасы үш-үштен 9 топқа топтасқан 27 микротүтікшелерден құрылған. Жасуша орталығының қызметі митоздың қалыпты жүруін қамтамасыз ету, яғни, анафаза кезінде хроматидалардын полюстерге ажырауын камтамасыз ету болып табылады.

Эндокринологияға кіріспе Жоғары сатыдағы жануарлар организмінің миллиарттаған клеткалардан тұратыны белгілі және ол бәрі бір-бірімен тығыз байланысқан. Осыншама көп клеткалар өзара үйлесімді әрекет жасайды, сондықтан да олар тірі организм денесі біртұтас болып табылады. Клеткалардың үйлесімді әрекетін нерв жүйесінде және эндокрин бездерінде жасалатын заттар реттеп отырады.
Нерв жүйесі нейрондардан, нерв клеткаларынан тұрады, оларда нейромедиатор заты түзіледі, нерв жүйесіндегі қоздыру, хабар беру осылар арқылы іске асады. Нерв жүйесі арқылы хабар беру өте тез - миллисекунд ішінде беріледі. Нерв клеткалары бірімен бірі өзара тікелей байланысқан.
Эндокрин (гректің endo krin- ішке шығарамын деген сөзінен шыққан) бездерінде немесе ішкі секреция бездерінде гормондар түзіледі, ол гормондар қанға қосылады да, қан арқылы синтезделген жерден қашықта орналасқан нысана - клеткаға жетеді. Эндокрин бездерінің гормон арқылы берілетін хабары тез емес, баяу. жетеді, бірақ оның әсері ұзақ болады.
Гормондардың түзілуін нерв жүйесі, әсіресе ми қабығы реттеп отырады. Адамның және жануарлардың бүкіл өмір бойы тіршілік әрекетін, организмдегі зат алмасуын орталық нерв жүйесі басқарып отырады.
Сонымен, гормон дегеніміз - биохимиялық процестерді реттейтін және адам мен жануарлардың аса маңызды тіршілік қызметіне қоздырушы әсер ететін химиялық зат.
«Гормон» деген термин гректің қоздырамын деген сөзінен шыққан (һогmао - қоздырамын, қозғалтамын), оны ғылыми ұғым ретінде 1904 жылы У. Бэйлисс пен Э. Стерлинг енгізген.
Гормондар тек нысана - клетка деп аталатындарға ғана әсер етеді, ол клеткалардың ерекше маманданған рецепторлары (rесерtоr - қабылдаушы деген латын сөзінен шыққан) болады, ондай рецепторлар плазмалық мембранаға немесе клетка ішіне орналасқан және олармен байланысқан. Гормондар тиісті рецепторлармен өзара әрекеттесу нәтижесінде ғана биохимиялық процестерге әсер етеді, биохимиялық процестерді тездетеді, ал кейде тежейді. Гормонның жарты өмірі ұзақ емес, әдетте ол бір сағаттан кем.
Гормондар организмде аса маңызды үш түрлі қызмет атқарады: 1) физикалық, жыныстық жетілуіне және ақыл-ойдың дамуына көмектеседі; 2) физиологиялық мүмкіндік жасайды және оны қамтамасыз етеді; 3) бірқатар аса маңызды физиологиялық көрсеткіштердің (осмостық қысымның, қандағы глюкоза деңгейінің) бірқалыпта болуын қамтамасыз етеді.
Ішкі секреция бездерінің, гипофиздің гормондары және гипо-таламустың гормрндары _жақсы зерттелген. Гипоталамус эндокрин жүйесін негізгі реттеуіш болып табылады-және мұнда <<өте күшті гормондар>> немесе <<нейрогормондар>> түзіледі. Олар басқа клеткаларда да гормонның түзілуіне әсер етеді.
Эндокрин бездерінің құрылымы мен қызметін, одан белініп шығатын секреттерді және зат алмасуды гормондардың реттеу механизмін зерттейтін биохимия тарауын эндокринология деп атайды.
Эндокринологияның негізін қалаған француз ғалымы Броун-Секар. Ол 1890 жылы жас қошқарлардың енінен алынған экстрактқа қатысты классикалық тәжірибе жасады. Ондай экстракты кәрі қошқарлардың денесіне (терісінің астына) құяды, оны бірнеше рет қайталағаннан кейін кәрі қошқарлар ширайды, олардың қимыл-әрекеті жігерлі бола бастайды, тіршілік қабілеті жақсарады, қан қысымы қалпына түседі, жемшөпке тәбеті артады. Осы тәжірибені өзіне істеп байқайды (ол 72 жаста еді) . Тәжірибеден кейін өзінде жаңа күш-қуат пайда болғанын сезіне-ді. Осындай тәжірибелерге сүйеніп, Броун-Секар мынадай қоры-тынды жасайды: жыныс бездері және басқа да бездер канға жігерлі заттар бөліп шығарады, ол заттар бүкіл организмге және жеке мүшелерге қосымша қуат береді.
Соңғы кездерде өсімдіктерден де олардың өсуін реттейтін гормондар табылды. Химиялық табиғатына қарай гориондар 4 топқа бөлінеді.
1. Белоктық және пептидтік: инсулин, глюкагон, гипофиз, гипоталамус, қалқанша маңындағы без гормондары.
2. Тироксин, адреналин, норадреналин, т. с. с. амин қышқыл-дарының туындылары болып табылатын гормондар.
3. Стероидтық: жыныс бездерінің гормоны, бүйрек безі гор-моны, т. б.
4. Жергілікті әсер ететін гормондар (гормон тәрізді заттар) : полиқанықпаған май қышқылдарының туындысы - простаглан-диндер.
ҚАЛҚАНША БЕЗІНіҢ ГОРМОНДАРЫ
Қалқанша без - ішкі секреция бездерінің ішіндегі негізгілерінің біреуі. Организмде айналып отыратын барлық қан тәулік бойы бірнеше рет қалқанша безі арқылы өтеді де, оның бөліп шығарған секретін барлық тканьдерге және мүшелерге жеткізе-ді. Қалқанша без гормондарыньің зат алмасуын реттеудегі маңызын осыдан-ақ байқауға болады.
Сүтқоректі жануарлардың қалқанша безі мойынңың ортасына таман, көмейдің астыиа орналасқан, оның пішіні қалқанға немесе тағаға ұқсас болады. Ол оншама үлкен емес. Ірі қараны-кі-15-45 г, жылқыныкі - 20-35 г, шошқаныкі - 12-30 г, қойдыкі - 4-7 г, адамдыкі-15-30 г шамасындай. Бұл без көптеген көпіршік клеткалардан (фолликулалардан) құралады, ол фолликулалар секретор эпителімен қоршалған, ішінде «кол-лоид» дея аталатын қоймалжың массасы болады. Қоллоид құра-мында белок, йодқа бай тиреоглобулин бар (м. м. 680 000) .
Қалқанша без организмге сумен және қоректік затпен бірге енетін йодидтерді өзіне шексіз сіңіреді.
X и м и я л ы қ т а б и ғ а т ы. Қалқанша бездің негізгі де әсі-ресе активті гормоны тироксин мен трнйодтиронин. Ол гормон-дар бірнеше рет айналып езгеру нәтижесінде тирозин амин қыш-қылынан түзіледі. Йодидтер ерекше пероксидаза ферментінін әсер етуімен тиреоглобулин белогы құрамындағы тирозин қалдығымен реакцияласады. Мұнда тирозиннің бүйірлік тобы йодтанады және монойодтирозиндер мен дийодтирозиндер түзіледі. Тиреоглобулин гидролизі кезінде тирозинйің йодтанған туындылары және басқа да амин қышқылдары босайды. Тироксин дийодтирозиннің екі молекуласының'тотыға конденсациялану жолымен түзіледі, бұл кезде аланин бәлініп шығады. Ал монойодтирозин мен дийодтирозиннен трийодтиронин түзіледі.
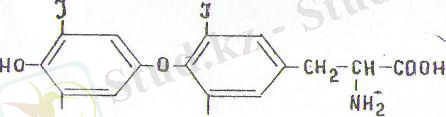
Тироксии (тетрайодтиронии)
Протеолитдік ферменттердің тиреоглобулин гидролизіне әсеріне және. осы бездің гормон бәліп, оны қанға жіберуіне типофиздің тиреотроп гормоны себепші болады және реттейді.
Зат алмасуға әсері. Тиреоидты гормондардың әсері ген аппараты арқылы іске асады. Олар хроматинде ядро ішіндегі белоктармен байланысады және ирНҚ транскрипциясын күшейтеді, жылдамдатады («Нуклеин қышқылдары» деген тарауды қараңыз) . Бұл жағдай белок синтезінің жылдамдығын және көптеген ферменттердің активтілігін арттырады.
Бұл гормондар іс жүзінде организмнің барлық органдары мен тканьдерінде биохимиялық реакцияларды тездетеді. Соның нәтижесінде белок алмасу, йод алмасу, углеводтардың алмасуы және тұздардың алмасуы тездейді, оттегін сіңіру және кемірқышқыл газды бөліп шығару артады. Осылардың нәтижесінде өсіп даму, организм тканьдерінің ыдырауы тездейді және орта-лық нерв жүйесінің жұмысына әсер етеді.
Қалқанша безінің жұмысы бұзылған кезде гормондардың бөлініп шығуы бәсеңдейді (гипофункция) немесе олар кептеп бөліне бастайды (гиперфункция) .
Гипофункция кезінде сүтқоректі жануарлардың зат алмасуы төмендейді. Мұндай жағдай жануарлардың кысқы ұйқысына және түлеуіне, оларға жаңа жүн шығуына себепші болады. Жануарлардың өнімділігі айтарлықтай төмендейді. Гормон шығуы азайған кезде адамдарда енжарлық, селқостық пайда болады, ұйқы басады, ойлау қабілеті төмендейді, ақылынан алжасады, Ол жағдай жас кезде кездессе, . өсуі тоқтайды (ергежейлі болып қалады), дене бітімі өзгереді. Медицинада мұны кретинизм деп атайды
Гипофункцияның басқа түрі - эндемиялық жемсау- (зоб) . Ондай сырқат су мен азық-түлікте йод аз болатын таулы аудан-дарда кездеседі. Бұл жағдай жануарлардың да денсаулығына және өнімділігіне жайсыз әсер етеді. Сондықтан ас тұзына йодты калийді қосып беру қажет.
Гиперфункция кезінде гормонның жасалып шығуы артады. Б. ұл кезде зат алмасу күшейеді, сырқат адамда қорқақтық пай-да болады, бойын үнемі қорқыныш билейді, нерв жүйесі қозьга, жүйкесі жұқарады, көзі ұясынан шығып, базед ауруына ұшы-райды. Сол сияқты мұндай кезде азот алмасуының кері құбылы-сына ұшырауы да мүмкін, бауырда гликоген өте азаяды, қоректі қанша көп жесе де организм жүдеп, денесі арықтайды.
ҚАЛҚАНША МАҢЫНДАҒЫ БЕЗДЕРДІҢ ГОРМОНДАРЫ
Сүтқоректі жануарларда қалқанша маңындағы безі жұптасқан екеу болады. Олар қалқанша бездің үстіңгі жағына орналасады.
Химиялық т а б и ғ а т ы. Қалқанша маңындағы бездерде екі пептидтік гормон - паратгормон (паратирин) және қальци-тонин түзіледі. Бұл екі гормон да бірқатар жануарлардың және адамның қалқанша маңындағы бездерінен таза күйінде бөлініп алынған. Олардың бірінші реттік құрылымы анықталған. Парат-гормон дегеніміз - полипептид.
Ол 83 амин қышқылы қалдықтарынан тұрады, м. м. 9500. Ал кальцитонин 32 амин қышқылы қалдықтарынан тұрады, м. м . 3600.
Зат алмасуға әсері. Екі гормон да Са 2 + мен фосфор алмасуына әсер етеді. Олар қандағы, ткань сұйығындағы және сүйек . тканіндегі кальций мен фосфор мөлшерін реттейді. Парат-гормонның нысана-мүшелері - суйек ткані мен бүйрек. Оларға паратгормон цАМФ («циклдік АМФ» қараңыз) арқылы әсер етеді.
Организмде паратгормон қалыпты мөлшерден артық болған кезде сүйек тканьдері бүлініп, бұзыла бастайды, ал гормон же-тіспеген кезде қанда және басқа да сұйық тканьдерде кальций иондарыныц мөлшері азаяды. Бұл кезде нерв жүйесінің қозғыш-тығы артады және кенеттен сіңір түйіліп, тарамыс тартылады.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz