Организмдегі көмірсулар алмасуы: қорытылу, сіңуі және гликоген метаболизмінің механизмдері

Организмдегі көмірсулардың алмасуы
Жоспар
- Зат алмасудың жалпы заңдылығы.
- Көмірсулардың қорытылуы.
- Күйіс қайтармайтын мал асқазанында көмірсулардың қорытылуы.
- Моносахаридтердің сіңуі.
- Күйіс қайыратын жануарлар мен жылқы организмінде көмірсулардың қорытылу ерекшелігі.
- Қан құрамындағы моносахаридтер.
- Гликогеннің бұлшық етте және бауырда синтезделуі мен ыдырауы.
- Көмірсу емес қосылыстардан көмірсулардың түзілуі.
- Фруктоза, галактоза және мальтозаның алмасуы.
- Гликолиз. Гликолиз реакцияларының реттілігі.
- Көмірсулар алмасуының реттелуі.
Зат алмасудың жалпы заңдылығы
Тірі организм дегеніміз-белгілі құрылымы бар биологиялық жүйе. Тірі организиде үздіксіз зат алмасу жүріп жатады. Зат алмасу дегеніміздің өзі организмде жүріп жататын барлық химиялық процестердің жиынтығы. Организмдегі зат алмасу тіршіліктің негізгі белгісі болып табылады. Тірі организм зат алмасудың нәтижесінде пайда болып жарыққа шығады, тіршілік етеді, өсіп жетіледі және өледі.
Соңғы кездерде әдебиеттерде « зат алмасу» дегеннің орнына «метаболизм» деген термин жиі қолданылатын болды бұл екеуіде бір мағына береді, сондықтан өзара бірін-бірі ауыстырады. Организмдегі зат алмасу (метаболизм) өзара тығыз байланысты екі процестен - катаболизм анаболизмнен құралады.
Катаболизм (гректің каtа- төмен, ballen- тастау деген сөздерінен шыққан ) дегеніміз күрделі органикалық қосылыстардың ыдырап, қарапайым молекулаларға айналуы. Азық - түлік, жем- шөп арқылы организмге енген және тканьдер мен органдарға барған белоктар, липиттер, көмірсулар бірқатар катаболиздік реакциялар нәтижесінде ыдырап, ақырғы өнімдерге айналады да, сыртқа бөлініп шығады. Катаболиздік реакциялар кезінде органикалық молекулалардан бос энергия бөлініп шығады және олар АТФ макроэнергиялық байланысында, басқада нуклеотидтерде жиналады. Жиналған осы энергия тірі организмнің тіршілік әрекетін іске асыру үшін жұмсалады.
Анаболизм (гректің аna-жоғары деген сөзінен шыққан) - бұл белоктар, липидтер, нуклеин қышқылдарының және клетка мен тканьдегі басқа да биомолекулалардың синтезі. Мұндай синтез катаболизм кезінде пайда болған заттардың есебінен іске асады. Анаболаизм реакциялары кезінде АТФ макроэргиялық байланыстарынан босап шыққан энергия жұмсалады. Анаболизм процесі кезінде организмнің құрам бөліктері қалыптасады және жаңа бөліктер түзіліп жаңарады. Ал организмді тұтастай алып қарайтын болсақ, ересек организмнің дене құрамы бірқатар уақыт бойы оншама өзгере қоймайтыны белгілі.
Клеткадағы катаболиздік және анаболиздік реакциялар бір-біріне тәуелсіз болады және бір мезгілде жүреді. Бұл екеуі зат алмасудың біртұтастығын және оның мазмұнын көрсетеді. Бұл реакцияларды ферменттер катализдейді және оны эндокриндік жүйе мен орталық нерв жүйесі басқарып реттейді. Катаболизм және анаболизм нәтижесінде түзілетін қосылыстар метаболиттер немесе аралық өнімдер деп аталады. Катаболизм диссимиляция деп, ал анаболизм ассимиляция деп те аталады.
Негізгі қоректік заттар (белоктар, көмірсулар, липидтер) - күрделі де жоғары молекулалы қосылыстар. Сондықтан олар ас қорыту жолында ішек-қарын қабырғасы арқылы сорылып, организмге ене алмайды. Ол заттар организмге сіңіп, оның құрам бөлігіне айналу үшін, оларды ыдырату және қарапайым молекулалы заттарға айналдыру қажет. Қоректік заттардың ыдырап айырылуы әр түрлі ферменттердің әсері арқылы іске асады және ол ас қорытылу процесі деп аталады. Ас қорытылу процесіне қатысатын ферменттер гидролазалар класына жатады. Осыған байланысты ас қорыту жолында ол ферменттер қатысатын катаболизм реакцияларындағы екінші субстрат су болады. Су минералдық тұздар және витаминдер де қоректік заттарға жатады.
Көмірсулардың қорытылуы
Көмірсулар жануарлар үшін негізгі қорек. Ол азық рационының 70% шамасындай. Ал адам қорегінің 50% углеводтардың үлесіне тиеді. Қорытылу ерекшелігіне қарай углеводтарды мынадай екі топқа бөледі: 1) крахмал, гликоген, сахароза, лактоза, 2) целлюлоза, маннандар, лигнин, пектиндер, пентозандар және басқалары. Екінші топқа жататын углевводтарды омыртқалылардың бәрі бірдей қорыта алмайды. Оларды күйіс қайыратын және кейбір жануарлар ғана микробтық ферменттің көмегімен қорыта алады (бұл мәселе жеке қарастырылады) .
Крахмалдың, гликогеннің қорытылуы ауыз қуысынан басталады, ол екеуінің қорытылыуна аздап болса да сілемей ферменттері әсер етеді. Сілемейдің негізгі қызметі - қоректі ылғалдандыру, сөйтіп оның шайналуына және ас қорыту жолымен жүруіне жағдай жасау. Сілемейдің құрамында, шырышты гликопротеидтер (муциндер) бар, олар қорекке жағылып, оны жұмсартады.
Сілемей дегеніміз түссіз (қойда, жылқыда) немесе азғана майлы су түсіндей (сиырда, итте) сұйық зат. Ірі қараның және қойдың сілемейінің рН көрсеткіші 8, 2-8, 5 жылқыда 7, 2-7, 8 адамда 6, 9-7, 0. Кейбір сүтқоректі жануарлардың сілемейі құрамында крахмал мен мальтозаны ыдырататын α -амилаза (м. м 56000) және мальтаза (м. м. 22000) ферменттері бар. Сілемей құрамында α-амилаза бейтарап ортада және шала қышқыл ортада активтілік қасиет көрсететіндіктен сілемей амилазасының активтілігі шамалы. Мысалы, адамдағы осы ферменттің активтілігімен салыстырғанда, шамамен шошқада 100 есе, ірі қарада, жылқыда 1400 есе кем болады.
Күйіс қайырмайтын мал асқазанында көмірсулардың
қорытылуы
Қоректік заттар ауыз қуысынан әрі қарай селінід күшті қыш-қылдық реакциясы (рН 1, 5-2, 5) бар қарынға түссді. Қышқыл ортада сілемейдегі а-амилаза ферменті өзініц осерін тоқтатады да, углеводтар бұдан әрі ыдырамайды. Қарын сөлінде амилаза ферменті болмайды. Крахмал, гликоген, сахароза, лактозаның қорытылуы негізінен он скі елі ішектс, ұйкы безі фермснттерінің және ішск шырыіпыныц әсерінеи іске асады.
Омыртқалы жануарлардыц ұйқы безінен шыгатыи сөлінде сілтілік реакциясы (рН - 7, 5-8, 5) бар көп мөлшерде натрий бикарбонаты болады. Ішектіц шырышынаи да натрий, бикарбо-нат, хлор иондары бөлініп шығады. Осының арқасында қарында бөлініп ніығатын тұз қышқылы бейтараптанады және ішектегі рН фермснттердіц асср стуінс сц қолайлы, 7, 5- 8, 5 децгейінде тұрақтандырылады.
Қоректіц негізгі бөлігі болып табылатын крахмал ауыз қуы-сында аздаи қана қорытылады, өйткені қорек ауыз қуысында ұзақ болмайды және жануарлар сілемсйініц рН көрсеткіші фер-менттер әсері үшін қолайлы емес.
Крахмал амилазадан және амилопектиннен құралады. Ол екеуішң үйқы безінен шыққан а-амилаза ферменті әсерінен бола-тын гидролизі ретсіз смсс, 2-~ 6 байланыстарды а(1->4) -байла-нысы бойынша біртіндеп ыдырату жолымен жүреді. Бұл глюко-за, мальтоза және ірі олигосахаридтер қоспасының пайда болуы-на океп соғады. Ал а-амилаза, ферментінің ос(1-»-6) -байланысын
гидролиздеу қабілеті жоқ. Сондықтан тек амилаза ғана жаңағы айтқан енімдерге бөлінеді. а(1-»-6) -байланысты тармақтары бар амилопектин (гликоген де) біршама ғана ыдырайды. Дегенмен амилопектин молекуласының тармақталғаи жеріндегі a(l-v6) -гликозидтік байланысын үзетін изомальтоза ферменті а(1->6) -гликозидаза] болады. Соның әсерінен изомальтоза тузіледі. Изо-мальтазаның әсері нәтижесінде амилопектин қалдыгы сызықтык құрылымға ие болады да, орі қарай ос-амилазамен гидролизде-неді.
Ішсктіц ү-амилаза фермснті олигосахаридтердің шетінен глю-коза қалдықтарын біртіндеп үзіп, ажыратады. Ал мальтаза фер-менті мен изомальтаза ферменті (м. м. 280 000) тиісіншс малъ-тозаны жоне изомальтозаны а-глюкозаныц скі молекуласына гидролиздейді.
Сонымен, крахмалды жануар мен адам организмдегі сілскей, амилаза фсрментініц әсері, ұйқы безінің сөлі мен ферменгтсрінің және ішек шырышыиың әсері нотижссінде сіңірсді. Осы аталған ферменттердіц біртіндеп әсер етуі нотижссінде крахмал ce-D-глю-козага дейін толық гидролизденеді. Бұдан кейін а-О-глюкоза ішектің эпиталий клеткаларының мембранасы арқылы сіцеді. Крахмал (гликогсн) гидролизініц жиынтық реакциясы төменде-гідей.
(СвНІоОв), , +/гН 2 О-«ІСвНІяОе
a-D-ғлюкоза
Сүттің негізгі углеводы дисахарид лактоза, сүтқоректі жа-нуарлардың ішек шырышындағы лактаза ферментініц әссрінён гидролиздснеді. Бұл кезде гликозидтік байланыс р(1->-4) үзіледі де, p-D-галактоза және a-D-глюкоза түзіледі.
СІ 2 Н2аО п + Н а О-»СвН Іа Об + С 6 НІ 8 Об
p-D-галактоза a-D-глюкоза
Жер бетінде тіршілік ететін сүтқоректі жануарлар төлдеу ал-дында жонс төлдегеннен ксйін, олардыц ішегіиіц бұдырлы жиек-шесінде көп мөлшсрде лактаза болады.
Қоректің ерекшелігіне байланысты сүтқорсктілсрдің ішегінде сахараза ферменті пайда болады, ол фермснт сахарозаны р(1->-->-2) -байланысы бойынша a-D-глюкозаға және p-D-фруктозаға ажыратып бөледі.
С І 2 Н 22 Оп + Н 2 О-^С 6 Н 12 Об + С 6 Н 12 Об
Сахароза a-D-глюкоза р-Ь-фруктозаМоносахаридтердің сіңуі
Ішек қабыргасы арқылы моносахаридтер ғана сіцеді, ал ди-сахаридтер мен күрделі углеводтардың мұндай қабілеті болман-ды. D-глюкоза мен D-галактоза концентрация градиснтіне қарсы активті тасымалдау жолымен эпителий клеткалары арқылы сі-ңеді. Активті түрде тасымалдау тек D-қатарындагы қанттарға ғана тән, фруктоза активті тасымалданбайды, ол араласу жолы-мен сорылады.
Қанттар ішектің саңылауынан клетка мембранасы аркылы таратқыш молекулалар кемегімен және натрий иондарының қа-тысуымен тасымалданады. Таратқыш-молекулада глюкоза мен Na+ ионыныц байланысатын жері бар. Ішектіц шырышты қабы-ғы шегінде таратқыш олармен байланысады да, клеткаға енеді. Клеткада олар бір-бірінен бөлінеді, глюкоза сероз мембранасы арқылы қанға барады, Na f шұғыл түрде тасымалданып кетеді, ал таратқьші-молскула қайтадан ішектіц қуысына келеді. Қант-тардың сіңу жылдамдығы ор түрлі. Галактоза мсн глюкоза тезі-рек сіңеді. Ал глюкозамен салыстырғанда фруктоза 2 есс, ман-ноза мен иентоза 5-10 ссе баяу сіцірілсді.
Күйіс қайыратын жануарлар мен жылңы организмінде көмірсулардың қорытылу ерекшелігі
Ac корыту жолыида қызмот атқаратьш бсздордс Ітолисаха-ридтердіц р-гликозидтік байланысын үзіІІ ажырататып формснг-тер сіпітсздслмсйді. ІІІөппеп қоректенстін жануарлар үшін р-гликозндтік баііланысы бар дсллюлоза жоне баска да күрдслі угловодтар пстізгі азық болып табылады. Бүл полпсахаридтер ас қорыту жолында тіріпілік стстіи микрооргапизмдсрдіц көме-гімеп қорытылады.
Күйіс қаныратым малдыц қарыны төрт бөлімнсн тұрады. Олар: үлксмі қарын, қатбаріпақ, жүмыріпақ л<оие үлтабар. Ал үлкоп қатбарпіақ жоне жұмыршақ қарында үнемі 1 мл қорсктік массада 10" бактериялар және 10 6 қарапайым жондіктер, ана-эробтық микроорганизмдер тіршілік етеді.
Жылқыныц тік іпюғі мен сокыр ііпсгіиің көлемі үлксн бола-ды. Жылқыныц осы ііпектерінде жәнс қүстардың бөтекесіндс көптоген микроорганнзмдсф тірпіілік отсді.
Мұндай жануарлардыц қарыныпда, ішсгінде тіршілік ететін микроорганизмдер клеткасында цсллюлаза, цсллобиаза фср-менттері жәнс басқа да фсрменттер түзілсді. Ондай фермснттер целлюлозаны, пектииді, цсллобиозаны гидролиздеп, нсгізінен р-глюкозага және ор түрлі моносахаридтерге айналдырады. Осы моносахаридтер және басқа да қарапайым канттар одан әрі қа-рай басқа микробтық ферменттердің осерінен ашиды да, нсгізінен сірке, пропион, сүт, май, пирожүзім, қымыздык, янтарь қышқылдары, басқа да органикалық қышқылдар, көмір қышқы-лы мен СН 4 түзіледі. Органикалық қышқылдар үлкен қарынның бсзсіз эпителийі арқылы сіцеді.
Полисахаридтер талшықтарының қорытылуы малдыц күйіс қайыру процесінде жеңілдейді. Күйіс қайыратын мал ірі шөпті шайнап ұсақтайды. Бүл кездс шеп ұнтағына ауа араласады, оны Іу; ал жұтып қарынға жібсреді. Лл шөппен бірге қарынға барған оттегі сірке қышқылдық ашуды жақсартады. Пайда болған сірке қышқылы майдыц түзілуіне жұмсалады. Сондықтан ірі азықтар сиыр сүтінің майлылығын арттыруға көмектеседі.
Күйіс қайыратын мал өте кәп мөлшерде сілекей бөліп (бір тәулікте сиырдың бөліп шығаратын сілекей мөлшері 100 л, қой-дыкі 10 л) шыгарады. Осы сілсксймен бірге үлкен қарынға, ішекке кеп мөлшерде бикарбонаттар мен мочевина барады. Бұ-лар ашу кезінде пайда болган қышқылдарды бейтараптайды. Көмір қышқыл газы (СО 2 ), сутегі (Н 2 ) жоие мстан (СН 4 ) сіц-бейді, кекіру кезінде сыртқа бөлініп шыгады, кемір қьпиқыл газыныц бір бөлігі басқа қосылыстар синтезделген ксздегі кар-боксилдену үшін жүмсалады.
Жогары сатыдагы өсімдіктср клеткасыныц қабығы нсгізінен цоллюлозадан құралады. Ал цсллюлозаның ыдырап бөлінуінде микробтық фермситтердің мацызы ерскше зор. Клстка ішіндегі корсктік заттар доллюлоза қабыгыиан қүтылып алған соц, ас қорытатын басқа фсрмснттсрдіц оссрінс үіпырайды да орғанизм-ге сіцеді.
Микроорганизмдср қышқыл реакциясы бар үлтабарға (нагыз қарынга) үздіксіз келіп жатады. Олардыц қорытылуы осы қа-рында басталады да, ішекке барып бітсді. Сонымсн күйіс кайы-ратын жануарлардың нағыз қорсгінің басым бөлігі өздерініц 1/7 массасыныц бөлігіндей болатын өз микрооргапизмдері екен.
Цоллголоза қи қалыптастыруіны көлсмдік масса рстінде және ішсктіц қабыргасына осср етстіп механикалық қоздырушы ретін-де адамга да, жаиуарларга да қажст. Цсллюлоза ішектің тол-қындана жиырылуын күшейтеді жәнс асқазан жолымен қоректің жылжып қозғалуына осер стеді.
Жануар (жыртқыштардан басқасы) цсллюлозасыз тіршілік ете алмайды. Үзақ уақыт бойы жануарларды целлюлозасыз қо-ректендірсс, ас қорытылуы бұзылады, тіпті елімге әкеліп соқты-рады.
Қан қүрамындағы моносахаридтер
Моносахаридтер мен органикалық қышқылдар ішектегі ка-пиллярлар арқылы қан тамырына барады да, қанмен бірге басқа мүшелерге, тканьдерге тарайды. Моносахаридтердің бір бөлігі - негізінен глюкоза, белгілібір мөлшерде ұдайы қан құрамында айналып жүреді. Оньщ деңгейі жануарлардың түріне байланыс-ты. Ол мына төмендегідей миллимоль өлшемімен берілген мәлі-меттерден (A. B. Чечеткин, 1982) көрінеді.
Ірі қара 2, 06 Тауық 7, 00
Шошқа 2, 80 Жылқы 3, 00
Ешкі 2, 38 Адам 3, 30-5, 30
Қой 3, 9 -
Ересек жануарлардың қанында глюкоза децгейі онша көп емес. Ішек арқылы гексозалардьщ шагын гана мөлшері сіцеді. Ал углеводтардыц негізгі бөлігінің сіңуіне, жемазықтың микроб-тық ферменттсрдіц осерінен сірке қышқылына және баска ұш-қыш қышқылдарға айналып, содан ксйін ғана қанға сіңетіндігі-не байланысты.
ГЛИКОГЕННІҢ БҰЛШЫҚ ЕТТЕ ЖӘНЕ БАУЫРДА СИНТЕЗДЕЛУІ МЕН ЫДЫРАУЫ
Гликогеннің синтезделуі
Клеткадағы углеводтар тотығу жоне энергия бөлу үшін ен тиімді субстрат. Бірақ клетка энергетикалық материал ретінде глюкозаны кор үшін жинай алмайды. Осіресе қацқа бұлшық ет-тсріндс және бауырда қор үшін жиналатын тиімді углевод - гликоген. Қандағы глюкозаның айтарлықтай бөлігі осы ткань-дерге жиналады да, гликогенге айналады. Бұл процесс гликоге-ногенез (гликоген түзілу) деп аталады. Гликогеногенез шагыи мөлшерде жануарлардыц барлык тканінде дерлік кездсседі.
Гликоген мөлшері шикі бауыр массасының 10% шамасындай, ірі қара бауырында 5% аспайды. Бұлшық сттегі гликоген мөл-шері аз, 0, 5% жуық. Жануарларды углеводтармен тойганша азықтандырганда бұлшық еттегі оның мөлшері 2% дсйін жстсді. Гликоген бауырдың беткі жагында түйіріпік ретінде жиналады, бірақ бауыр жұмысын бұзбайды.
Жануарлар аш болганда бауырдағы гликоген тез шығыи бо-ла бастайды да, 24 сагат ішінде мүлдс таусылады. Ал углевод-тарға бай азықтармен, әсіресе глюкозамен қоректендіргси кезде гликоген қалпына келеді. Қалыпты жағдайда бауырда жонс бұл-шық еттерде гликоген синтезі жоне оның ыдырауы үнемі болып жатады. Жануарлар ашыққан ксзде бұлшық ет тканінде глико-ген бауырдағыдай тез таусылмайды.
Бүлшық еттегі және бауырдағы гликогеногенез процесі бірдей жолмен, глюкозо-6-фосфат және УДФ-глюкоза арқылы жүреді деп топшылайды. Өйткені, бұл екі тканьде де бірдей ферменттер
табылған. Дегенмен, бауыр ферменттері мен бұлшық ет фер-менттері құрылымында және олардың активтілігін реттеу меха-низмінде айырмашылық бар. Мұндай ферменттік ерекшеліктерге тереңдемей, бүлшық еттерде және бауырда гликогеннің түзілуін қысқаша қарастырамыз. 1, Гликогеннің синтезделуі АТФ энергиясы есебінен бос глю-козаның фосфорлануы арқылы басталады, бүл реакцияны гексо-киназа ферменті (м. м. 104000) катализдейді, бұл фермент бар-лық клеткада кездессді. Осы реакцияны катализдейтін екінші фермент глюкокиназа (м. м. 2 000 000), бұл фсрмент организм туғаннан кейін скі жеті өтісімен пайда болады. Глюкозаныц фос-форлану реакциясы магний ионы қатысында глюкозо-6-фосфат түзілу арқылы жүреді. Глюкозо-6-фосфат - гликогснніц метабо-лизмі кезіндегі негізгі заттьщ бірсуі.
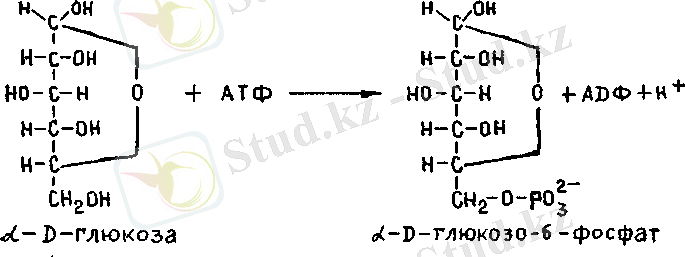
2. Фосфоглюкомутаза ферментінің (м. м. 62000) осерімен
глюкозо-6-фосфат қайтымды изомерлеиіп глюкозо-1-фосфатқа
айналады.
Глюкозо-6-фосфат ч=4= глюкозо-1-фосфат
3. Гликогсн синтезіндегі аса мацызды кезен уридинтрифос-
фаттың (УТФ) қатысуымсн глюкозаныц активтенуі жоне глюко-
заньщ активті формасы •-- (УДФ-
глюкоза) түзілуі:
Глюкозо-1 -фосфат + УТФ -> УДФ-глюкоза + пирофосфат. Бұл рсакщшны глюкозо-І- фер-мёнті каталнздсйді.
4. Гликоген синтезінің қорытынды кезеңінде гликоген-синте-
таза деп аталатын фермент реакцияға кіріседі, ол фермент глю-
козаны УДФ-глюкозадан шығарып, полисахаридтердің өсіп бара
жатқан тізбсгіне қосады. Гликогеногенездің бұл сатысы өте күр-
делі және оның механизмі негізінен анықталған. Біз оны схема
түрінде ғана берсміз.
УДФ-глюкоза гликоген соцындағы көміртегінің тертінші ато-
мыныц гидроксилдік тобына ауысады да, гликозидтік байланыс а(1->1) түзеді.
Егер олигосахаридтік тізбектс глюкоза қалдығы төртеуден көп болса ғана мұндай алмасу іске асады. Осыдан гликоген "син-тетазасы басқа синтетаза түзетін уытты қажет етеді. Бірақ клет-кадағы бұл уыттың шыққан тегі қазірше анықталған жоқ.
Гликоген молекуласының тармақталған екені белгілі, ондай тармақтар негізінен гликозидтік байланыс - а(1-»-6) адқылы жалғасқан. Гликоген-синтетаза тек а(1-»-4) байланысыныц пай-да болуын ғана катализдейді. Гликоген молекуласыньщ тармақ-талу нүктесінде а(1-»-6) -байланыс қалыптасу үшін гликозил - (4-6) - трансфераза ферменті қажет. Ол фермент глюкозаның 6-7 қалдығын гликогеннің С 6 гидроксильдік тобына ауысуын катализдейді. Осыдан кейін гликоген-синтетаза ферменті поли-сахарид тізбегін гликогсннің толық молекуласы түзілгенге дейін соза алады.
Гликоген синтезделуінің соңғы кезеңін қарапайым түрде бы-лай көрсетуге болады.
«УДФ-глюкоза->- (СбН 10 О 5 ) и + п(УДФ)
Гликоген молекуласы тармақталуының бағалы биологиялық мацызы бар. Ол полисахаридтердің ерігіштігін арттырады және оган катаболизм ферменттерінід әссріи күшсйтеді.
ГликогеІІ синтезін инсулин гормоиы стимулдсйді, инсулнн гликогеногснездіц бірқатар фсрменттсрінің активтенуінс, солар-дыц ішінде глнкогсн-синтетаза фсрментініц активтенуіне қаты-сады.
Гликогеннің ыдырауы
Гликогеннің маңызды қасистініц бірі - оның организмніц қа-жет отуіне қарай қор рстінде жиналатындығы жоне ыдырап глюкозага айнала алатындығы. Гликогенніц ыдырау процссі гли-когенолиз деп аталады. Қан құрамындагы глюкоза мөліпері тө-мендеген ксзде адреналин жоне глюкагон гормондарынын әсері-нен цАМФ синтсзі күшейоді де гликогенолиз процесі жүреді. Гликогенніц бұлшық отте көбірск ыдырауын адрсналин іске асырады, глюкагон гормоны гликогеішің бауырда ыдырауына күштірск осср етеді.
Гликогеннің бұлшық сттерде ыдырауының бауырда ыдырауы-нан айтарлықтай айырмасы бар және глюкозо-6-фосфат түзіл-геннен кейін ор түрлі жолмен жүреді. Бауырда глюкозо-6-фосфат гидролизденеді де, бос глюкозага және минералдық (органика-лық емес) фосфатқа бөлінеді. Бұдан кейін глюкоза қанға қосы-лады. Қанмен бірге ец алдымен қацқа бұлшық еттерінс жоне миға барады. Оның үздіксіз келіп тұруы бұл органдар үшін аса
қажет. Бұлшық етте гликоген глюкозо-6-фосфат арқылы пиру-ватқа дейін тотығады. Пируват бұдан кейін тотықсызданып, лак-татқа айналуы мүмкін. Бұл процесс гликолиз деп аталады, ол кейінірек қарастырылады. Енді гликогеннің бауырда глюкозаға айналуын қарастырайық.
Гликогенюлиз мынадай кезецдердсн тұрады:
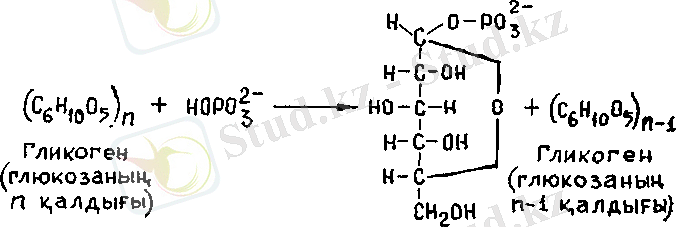
1. Гликогеннің ыдырау реакциясы фосфоролизден басталады, бұл кезде гликогенніц шеткі глюкозасы фосфоролиздік жолмен (гидролиздік емес) ажырайды да, оған бейорганикалық фосфат қосылады. Бұл процесті гликоген-фосфорилаза ферменті ката-лиздейді жоне глюкозо-1-фосфат түзіледі.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz