Эукариоттар мен прокариоттардағы ДНҚ зақымдануы және біртізбекті мен қостізбекті үзілістердің молекулалық репарация механизмдері

Кіріспе
Клетканың генетикалық ақпаратының тұтастығы ДНҚ бұзылысының кең спектрін шақыратын экзогендік химиялық және физикалық факторлар ықпалына ұшырап отырады. Оның үстіне, ДНҚ - ң спонтанды (белгілі бір бағытталған әсерлерсіз) зақымданулары қалыпты клеткалық метаболизм процесінде де пайда болады. Спонтанды немесе иондаушы радиация, метилметансульфонат алкилдеуші агенті сияқты физикалық және химиялық факторлардың әсерінен пайда болған ДНҚ-ң қостізбекті үзілістері ДНҚ репликациясы мен транскрипциясына кедергі болатын өте ауыр ДНҚ зақымданулары болып табылады. Репарирленбеген немесе дұрыс репарирленген қостізбекті ДНҚ үзілістері хромосомалық қайта құруларға, мутагенезге, канцерогенезге және клетканың өлуіне әкеледі. Барлық организмдердің маңызды қажеттілігі негом тұтастығын қолдау және ДНҚ зақымдарының қорлануына қарсы әрекет жасау болып табылады. Тірі организмдер осындай ДНҚ зақымдарын түзетуге арналған арнайы жүйелерді, арасында маңыздылары болып табылатын УК зақымдалудың фотореактивациясы, нуклеотидтер мен негіздердің эксцизионды репарациясы және ДНҚ-ның рекомбинациялық репарациясын жасап шығарды. ДНҚ-ның қостізбекті үзілістері көбінесе рекомбинативтік репарация жүйесі көмегімен ашытқы клеткаларында репарирленеді. Рекомбинациялық репарация ДНҚ қателіктерін түзетудің қателіксіз тәсілі болып табылады, өйткені басқа гомологтың немесе еншілес хроматидтің ақпаратын қолданады. Клеткада ДНҚ-ның қостізбекті үзілістерінің үзіліс орнындағы ДНҚ соңдарының гомологты емес бірігуі мен біртізбекті аймақтарының әлсіреуіне негізледген басқа да репарация механизмдері бар. Алайда рекомбинациялық репарациядан өзгешелігі қостізбекті ДНҚ үзілістерінің соңғы екі репарация механизмі мутагенді болып келеді, сондықтан олар үзіліс орындарында ДНҚ делециясын алып жүреді.
ДНҚ зақымдануына жауап ретінде клеткалық циклдің тоқтауы болады және ДНҚ репарациясының әртүрлі жолдары қосылады. Бұл процесстер ДНҚ зақымдары репарацияланбайынша митозға немесе S-фазаға клетканың кіруін болдырмайтын клеткалық циклдің бақылау механизмдерімен бақыланады. Клеткалық циклдің бақылау механизмдері ДНҚ зақымдалуына клеткалық жауаптың маңызды бөлігі болып табылады, және клетканың өмір сүруін және оның геномының тұрақтылығын қамтамасыз етеді.
Иондаушы радиация медициналық диагностикалық мақсаттарда және рак терапиясында құрал ретінде кеңінен қолданылатындықтан, ДНҚ - ның мүмкін зақымдануларына клеткалық жауаптың өте толық түсінігі қажет. Көбінесе рак клеткаларының радиациялық тұрақтылығы ракқа қарсы терапиямен мәселелер тудырады. Зерттеу бойынша, радиациялық тұрақтылық рак клеткаларының жоғары репарациялық клеткаларымен шақырылуы мүмкін. Қатерлі ісікке бейім осындай синдромы бар адамдардағы рекомбинацияның жоғары дәрежесі пациент клеткасында геномдық тұрақсыздықты шақырады. Сүт немесе аналық безінің қатерлі ісігіне бейімділіктің арасындағы тікелей байланыстың бар болуы және репарациондық процесстің бар болуы Rad51 басты рекомбинациялық белогымен Brca1 және Brca2 белоктарының өзара әсерлесуінің анықталуы жақында көрсетіліп шығарылды. Рекомбинациялық репарация эволюциялық кертартпа механизм болып табылады және бактериялардағы сияқты адамда да клетканың тірі қалуы мен геномның амандығының сақталуында шешуші рөлді ойнайды.
Осындай жолмен, ДНҚ-ның қостізбекті үзілістерін репарациялаудың молекулярлық механизмінің оқытылуы молекулярлы генетика мен биологияның өзекті мәселесі болып табылады. ДНҚ рекомбинациясы мен репарациясы механизмдеріне қатысатын жаңа гендердің ашылуы мен оқытылуы бұл процесстерді тереңірек түсінуге ғана емес, онкология мен генотерапия үшін практикалық мәнін де жете түсінуге мүмкіндік береді.
Курстық жұмыстың мақсаттары
Курстық жұмыстың басты мақсаты - эукариоттар мен прокариоттардағы ДНҚ-ның кез келген бұзылыстарының молекулярлық механизмдерін анықтау болып табылады. Сонымен қоса біртізбекті және қостізбекті ДНҚ үзілістерінің репарациясын зерттеу.
Зерттеу мақсаттарына сәйкес келесі міндеттер қойылды:
- ДНҚ-ның бұзылуына алып келетін экзогендік және эндогендік факторлардың әсерін анықтау.
- ДНҚ-ның зақымдалу түрлері мен типтерін жете танып білу.
- Олардың қателіктерін болдырмау шараларын жасау.
- Олардың тіпті қалыпты метаболизм кезінде де пайда болу жиілігін анықтау.
- Және бұл типтердің организмдердің әрқайсысына қандай дәрежеде әсер ететінін және олардың клеткасында қалай іске асатынын анықтау (репарацияның әр түрі прокариоттар мен эукариоттарда әртүрлі іске асады) .
- Репарацияны жүзеге асыру кезінде жаңа гендерді ашып, олардың белоктармен өзара байланысын ашып көрсету.
- Осы репарацияның типтерінің ДНҚ-ның зақымдануын түзету кезіндегі пайда болған қателіктерінің нәтижесінде олардың қандай генетикалық ауруларға ұшырататынын анықтап көрсету және оларды сипаттау.
1. ДНҚ репарациясына қысқаша анықтама
Репарация - тірі ағзалардың мутагендік факторлар әрекетіне байланысты ДНК молекуласында пайда болған бұзылуларды қайта қалпына келтіру қасиеті. Латынша “ reparatio” - түзелу, қалпына келу деген мағынаны береді. Репарация құбылысы тек ДНК құрылысындағы мутациялық бұзылуларды жөндеуде емес, сонымен қатар тұқым қуалайтын аурулардың қалыптасуында, ерте қартаю процесінде, жасушалардың ісіктік трансформациясында (канцерогенез) т. б. маңызды рөл атқарады. Репарация бір жасушалыларда, өсімдік жасушаларында және жануарларда тірі қалу деңгейін көтеріп, аберациямен гендік мутацияларды төмендетеді. Репарация ерекше ферметтер арқылы жүзеге асады.
ДНҚ репликациясы РНҚ мен белок синтезі сияқты процестерге қарағанда әр клеткада әр ұрпақ сайын тек бір рет қана іске асады. Негізінде, репликация процесінің нақтылығы жоғары болуы және мутацияларды болдыртпау өте маңызды. Мутациялар репликация кезінде пайда болатын қателердің нәтижесінде орын алады және олардың кейбір түрлері зиянды, кейде қазалы болуы мүмкін. Табиғат ДНҚ тізбегінің нақты түрде көшірілуін қамтамасыз ететін бірнеше жолын жетілдірді. Жалпы, мутациялық өзгергіштік гендер мен хромосомалардағы тұрақты өзгерістер нәтижесінде қалыптасады да, генетикалық материалдың геномдық, хромосомалық және гендік (ДНК молекуласы) деңгейлеріндегі кез - келген сандық және құрылыстық бұзылуларға әкеледі. Осыған байланысты, яғни бұзылуларды қайта қалпына келтіру үшін ДНК репарациясы жұмыс істейді. ДНҚ - ның репарациялану мүмкіндігі 1949 жылы белгілі болды. Репликация қателері спонтанды түрде әр 10 9 -10 10 негіз сайын орын алады. Түзету механизмі репликация процесі кезінде дұрыс емес нуклеотидті тізбекке қосқаннан кейін лезде алып тастауға сүйенеді. ДНҚ-полимеразаның үш белсенді сайты болады. Сутектік байланыстағы қателіктер арқасында дұрыс емес нуклеотидтің енуі әр 10 4 -10 5 жұп нуклеотид сайын орын алады. ДНҚ-полимераза I дұрыс емес нуклеотидті өзінің 3’-экзонуклеазалық белсенділігінің көмегімен жояды. Дұрыс нуклеотид енгеннен кейін репликация жалғасады.
1. 1 ДНҚ репарациясына қатысатын ферменттердің рөлі
Репликация кезінде ДНҚ-полимераза I ферменті кесу-жамау процесін іске асырады. Кесу - бұл РНҚ праймерді 5’-экзонуклеазалық белсенділік көмегімен алып тастау, ал жамау - ферменттің полимеразалық қасиетінің көмегімен дезоксинуклеотидті тізбек құрамына енгізу болып табылады. Процестің бұл бөлігі ДНҚ-полимераза III ферментінің жаңа полинуклеотидтік тізбекті синтездегеннен кейін орын алады. Матрицалық ДНҚ-да полимераза I өзінің 5’→3’ экзонуклеазалық қасиетінің көмегімен РНҚ праймерді алып тастау және ДНҚ-дағы қателерді түзей алады. Осыдан кейін ол өзінің полимеразалық белсенділігінің көмегімен бос аймақтарды толтырады. Бұл процесс ник трансляция деп аталады. Полимераза I жалғыз полипептидтік тізбектен тұрады, ал полимераза II мен III ортақ мультисуббөлікті болып келеді және олардың кейбір суббөліктері ортақ болады. Полимераза II репликация үшін керек емес; ол түзететін репарация ферменті болып табылады. Жақында тағы екі полимераза : Pol IV және Pol V анықталды. Олар да түзететін ферменттерге жатады және клеткадағы SOS жауап атты уникальды түзету механизміне қатысады. Полимеразалардың көңіл аударатын екі қасиеті бар: синтездеу реакциясының жылдамдығы (айналым саны) мен процессивтілік (фермент матрицадан диссоциацияланғанға дейінгі уақыт аралығында байланысқан нуклеотидтер саны) болып табылады. Қазіргі таңда ДНҚ-полимераза I ферментінің репарация мен түзетуге, ал ДНҚ-полимераза III ферментінің полинуклеотид синтезіне жауапты екені белгілі. ДНҚ-полимераза II, IV және V ферменттерінің функциясы - репарация болып табылады. Экзонуклеазалық белсенділік - ДНҚ полимеразаның түзету және репарация қасиеті үшін қажет. Осы процесс кезінде дұрыс емес нуклеотидтер жойылып, керекті нуклеотид тізбекке енгізіледі. Барлық үш ферментке ортақ болатын 3’→5’ бағыттағы экзонуклеазалық белсенділік түзету функциясына қажет; дұрыс емес нуклеотид синтез барысында жойылады да, дұрыс нуклеотид тізбек құрамына енгізіледі. Түзету процесі кезінде бір мезгілде тек бір нуклеотид түзетіледі. 5’→3’ бағыттағы экзонуклеазалық белсенділік көмегімен репарация кезінде нуклеотидтердің қысқа тізбегі жойылады. Кейбір ферменттердің түзету мен репарация қасиетінің эффективтілігі төмен болады.
Репарацияның биологиялық мәні: ол генетикалық материалдың мұрагерлік бұзылысы - мутацияның түзілуінің алдын ала отырып, ДНҚ молекуласындағы зақымдалуларды жояды. Шамалап айтқанда әр 9 секунд сайын ДНҚ тіршілік әрекеті кезінде зақымдалады. Зақымдалған клеткада ол өлу мүмкін болмаса, зақымдардың қайсысы болса да тез жойылады. ДНҚ репарациясына 150 ден аса гендер қатысады[2] .
1. 2 ДНҚ-ның зақымдалу типтері
Зақымдалған ДНҚ мынадай типтерге бөлінеді:
1. Репарацияланатын және репарацияланбайтын;
2. Спонтанды және индуцияланатын;
3. Индуцияланатын эндогенді факторлар;
4. Индуцияланатын экзогенді факторлар.
1. Репарацияланатын зақымданулар клетканың өз жүйесімен жойылады, мысалы, УК сәуленің әсерінен пайда болатындар. ДНҚ зақымдарының басым көпшілігі репарацияланады. Репарацияланбайтын зақымданулар сирек кездеседі.
2. Спонтанды зақымдалулар белгілі бір бағытталған әсерлерсіз пайда болады. Ал индуцияланатындар физикалық, химиялық немесе биологиялық факторлардың әсерінен пайда болады. Күн сайын адамның әр клеткасында азоттық негіздер 2-ден 3 мыңға дейін пуриндік және пиримидиндік нуклеотидтерін жоғалтады. Нәтижесінде, АП-сайттар түзіледі(апуринді және апиримидинді) . Тек дезоксирибоза мен фосфодиэфирлік байланыстар сақталады. Спонтанды зақымдалуларға, сондай-ақ, азоттық негіздердің дезаминделуі де жатады. Күн сайын адам клеткасында шамамен 200 цитозин урацилге айналады. Оның үстіне, аденин гипоксантинге, гуанин ксантинге айналады. Метильді СН тобының оттегіне қосылуы цитозинді тиминге айналдырады. Күн сайын адамда 50 мың біртізбекті үзілістер, 8 мыңнан аса тотыққан және алкилденген негіздер және тағы біріккен 100 күрделі зақымдалулар (қостізбекті үзілістер, ДНҚ - ДНҚ, ДНҚ - ақуыздың молекулааралық коваленттік тігістері) пайда болып отырады. Әртүрлі типті ДНҚ - ның 1000 зақымдалуларының ішіндегі репарация жүйесінің арқасында тек біреуі ғана мутацияға алып келеді.
3. Индуцияланған зақымдалулар мыналарды тудырады:
Физикалық факторлар - радиацияның барлық түрлері, ультракүлгін сәуле, СВЧ, температура.
Химиялық факторлар - полициклді және гетероциклді ароматты көмірсулар, ароматтық аминдер, мутагендер (нитрозогуанидин мен этилметансульфонат), уретан, формальдегид, азотқышқылы.
Биологиялық факторлар - афлатоксин мен басқа эндо- және экзотоксиндер, оттектің активті формалары[9] .
Сурет-1-ДНҚ-ның зақымдалу түрлері

ДНҚ-ның жиі пайда болатын бұзылыстарына мыналарды жатқызуға болады:
- температураның өсуіне ұшырап отыратын пурин мен
дезоксирибоза арасындағы гликозидтік байланыстардың ажырауы (депуринизация) ;
- цитозин мен адениннің қалдықтарының спонтанды дезаминденуі;
- ерекше химиялық заттардың әсер етуімен (алкилдеуші агенттермен) азотты негіздердің алкилденуі;
- ультракүлгін сәулелерді жұту кезінде шынжырдағы көрші пиримидиндер арасында циклобутандық димерлердің түзілуі.
ДНҚ-да жиі әрі спонтанды түрде апуринизация мен негіздердің дезаминденуі жүреді. Апуринизацияның нәтижесі АП-сайт - негізінен айырылған дезоксирибоза болып табылады.
Сурет-2-Апуринизация
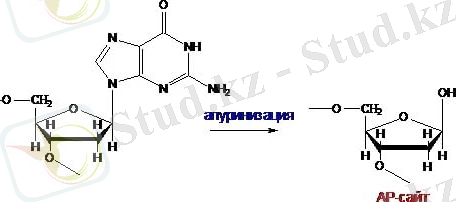
Цитозинді дезаминдеу кезінде ол урацилге, аденин - гипоксантинге, ал гуанин - ксантинге айналады.
Сурет-3-Цитозиннің дезаминделуі
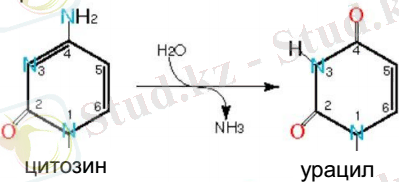
Атап өтілген бұзылыстардың көпшілігі репликация мен гендер экспрессиясының процестерін бұзады, мысалы, E. coli ДНҚ-сыдағы әр тиминдік димер репликацияны 10 секундқа кідіртеді. Оның үстіне, олар ДНҚ репликациясының басталуына дейін түзелмесе, бұл бұзылыстар мутацияға алып келеді. Көбінесе, мұндай бұзылыстар тек бір ДНҚ тізбегінде ғана болады, есесіне зақымданудың қарама-қарсы тұрған екінші тізбегінде көп жағдайда қателіктерді түзету үшін матрица арқылы қызмет ете алатын дұрыс бірізділік қалыптасады.
Мұндай тәсілмен ДНҚ-ның қос спиралі қателікті түзеу механизмін - репарацияны мүмкін етеді. Әртүрлі ағзаларда болатын репаративтік жүйелер мен механизмдер өте көп, олардың үшінде тек бір түрдің зақымдалуын түзеуге тән түрлері де бар.
2. ДНҚ репарациясының түрлері
Репаративтік механизмдерді тағы екі түрге бөлуге болады: жарық және қараңғы репарация.
Репарация типтерінің про- және эукариоттарда таралуы:
Прокариоттарда:
- Фотореактивация;
- Эксцизионды репарация;
- Пострепликациялық репарация;
- Қателікке бейімді репарация;
- SOS-репарация.
Эукариоттарда:
- Пострепликациялық репарация;
- Қателікке бейімді репарация;
- Қате жұпталған нуклеотидтердің репарациясы (Мисмэтч) ;
- Бір және тізбектік үзілістердің репарациясы.
Бактерияларда кем дегенде репарация жүргізетін 3 ферменттік жүйе бар:
- Тікелей;
- Эксцизионды;
- Репликациядан кейінгі.
Тікелей репарация - нуклеотидтердің бастапқы құрылысын қайта қалпына келтіре отырып, зақымдалуды тез арада жоюға қабілетті ерекше ферменттер әрекет ететін, ДНҚ зақымдалуын жоюдың ең оңай жолы.
Тікелей репарацияның түрлері:
- ДНҚ фотореактивациясының ферментативтік жүйесі;
- метилтрансферазаның қатысуымен метилденген гуаниннің репарациясы;
- лигазаның ДНҚ-полинуклеотидінің қатысуымен ДНҚ-ның бір тізбектік үзілуінің репарациясы;
- АП-сайттардың инсертаздармен репарациясы.
Зақымдалуды түзеудің тікелей репарациондық механизміне көрші пиримидиндік негіздердің арасындағы циклобутандық димерлердің мономеризациясы мен гуанин қалдықтарын дезалкилдеуді жатқызуға болады. Метилгуанин қалдықтарын дезалкилдеу қараңғы репарацияға жатады және сүтқоректілер мен бактериялардың клеткасына жататын ферменттер қатысымен жүреді. О 6 -метилгуанин-ДНҚ-алкилтрансфераза алкильдік топтардың ферментерінің цистеиндік қалдықтарының сульфгидрильдік топтарына тасымалын катализдейді.
Сурет-4-ДНҚ-алкилтрансферазамен О 6 -метилгуанин қалдықтарының дезалкилденуі
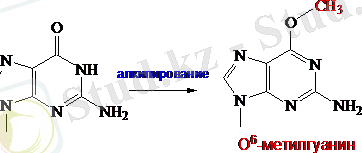
E. coliклеткасында минутына метилтрансферазаның 100 молекуласы синтезделуі мүмкін. Метилтрансферазаның мысалдары: О 6 -метил-гуанин-трансфераза (бактерияларда), О 4 -метил-тимин-ДНҚ-метилтрансфераза мен О 6 -метилгуанин-ДНҚ-метилтрансфераза (адамдарда) .
2. 1 Фотореактивация
Генетикалық кодты дұрыс оқымау нәтижесінде орын алатын спонтанды мутациялардан басқа организмдер мутацияның себебі болатын мутагендердің әсеріне тап болады. Кең таралған мутагендер қатарына ультракүлгін сәулелер, иондық радиация және көптеген химиялық агенттер жатады. Ультракүлгін сәулелерінің әсерінен ең жиі орын алатын мутацияларға пиримидин димерлерлерінің қалыптасуын жатқызуға болады.
Сурет-5-Циклобутил сақинасының түзілуі
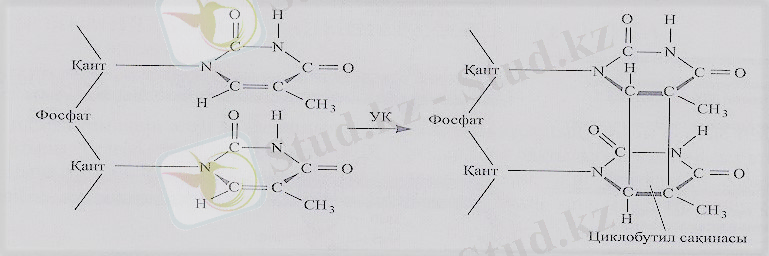
Екі жақын орналасқан пиримидиндердің екі π электроны циклобутил сақинасын түзеді де, ДНҚ-ның қалыпты пішінін бұзады. Әдетте бос радикалдармен келтірілген химиялық зақым нәтижесінде ДНҚ тізбегінің фосфодиэфирлік қаңқасын бұза алады. Бұл жағдай антиоксиданттардың соншама танымал екенінің себебі болып табылады.
Мутациялық өзгергіштік гендер мен хромосомалардағы тұрақты өзгерістер нәтижесінде қалыптасады да, генетикалық материалдың геномдық, хромосомалық және гендік (ДНК молекуласы) деңгейлеріндегі кез - келген сандық және құрылыстық бұзылуларға әкеледі. Осыған байланысты, яғни бұзылуларды қайта қалпына келтіру үшін ДНК репарациясы жұмыс істейді. ДНҚ - ның репарациялану мүмкіндігі 1949 жылы белгілі болды. А. Кольнер, Р. Дюлбекко және И. Ф. Ковалев бір - біріне байланыссыз түрде актиномициттер, бактериофаг және парамецияларға ультракүлгін сәулелердің летальды дозасымен әсер еткеннен кейін кәдімгі жарық сәулесін түсірсе олардың тіршілік қабілетінің қалпына келетіндігін анықтады. Бұл құбылыс фотореактивация деп аталады[1] .
Пиримидиндік нуклеотидтердің арасындағы димерлердің ыдырауы фотореактивация процесінде жүреді. Фотореактивация - көрінетін жарықтың әсер етуі нәтижесінде УК-сәулелермен зақымданған ДНҚ молекуласының құрылымының қалпына келуі. Тимин димерлері ДНК-ң құрылымын бұзу нәтижесінде ДНК реплекацияның өтуіне қиыншылық туады. Көк-күлгін жарық дезоксирибо-пиримидинфотолиаза ферментін белсендіреді. Нәтижесінде тимин димерлері бір-бірінен ажырап А-Т аралығы сутектік байланыс қалпына келеді. Оның ферменттік және ферменттік емес түрі белгілі. Ферменттік фотореактивация көрінетін жарықтың 300-600 нм ұзындықтағы толқынының қатысуын талап етеді және ерекше фотореактивациялық ферменттердің ( дезоксирибопиримидин- фотолиаза ) әсер етуімен жүзеге асады. Фотолиаза ферменті тиминдік димерлерді танып, олармен жарықта да, қараңғыда да кешен түзеді. Көрінетін жарықпен жарықтандыру кезінде фермент активациясы жүреді, циклобутан сақинасы үзіледі, және қайта екі тимин түзіледі. Фотолиазаның субстраты болып пиримидинік негіздер қызмет етеді(зақымдалмаған ДНҚ-мен фермент байланыспайды) . Жұтылған жарық энергиясын қолдана отырып, фермент ДНҚ тізбектерін ажыратпай-ақ димерді бұзады. Фотолиаза 1 минутта 204 ыдыратуға қабілетті. E. coli бактериясында фотореактивация жүйесі 90% пиримидиндік димерді жояды және тек бір генмен phr ғана бақыланады. Бұл генмен мутация әкелетін штаммдар ДНҚ-ны репарациялауға қабілетті емес. Фотореактивацияны тек циклобутандық димерлер ғана мүмкін етеді. Бұл активация факторы болып химиялық энергия емес, көрінетін жарық энергиясы қызмет ететін жалғыз ферменттік реакция екенін атап өту керек. Фермент фотон жарығын (көк жарықты) жұтады, сонан кейін пиримидиндік нуклеотидтер арасындағы циклобутандық нуклеотидтерді ыдырату қабілетін иеленеді.
Көрінетін жарық әсерінен кейін ультракүлгін сәулелердің летальды дозасымен емделген организмнің тіршілікке қабілеттігінің жоғарылауы көрсетілді. Фотореактивацияның эффектілігі рН деңгейіне, температура мен клетканың физиологиялық жағдайына байланысты. Фотореактивациякейбір балықтардың, құстардың, амфибияның, жәндіктердің, жоғарғы сатыдағы өсімдіктер мен балдырларда табылған. 1969 жылы фотореактивацияға қабілеттілікті қалталы жануарлар да иеленетіні дәлелденді. Соңғы жылдардағы зерттеулер адам терісі клеткаларында да фотореактивациялаушы ферменттердің барын көрсетеді. Қазіргі уақытта фотолиазаның Micrococcus бактериясынан басқа барлық организмдерде бар екені белгілі болды.
Сурет-6-Фотореактивация құбылысы
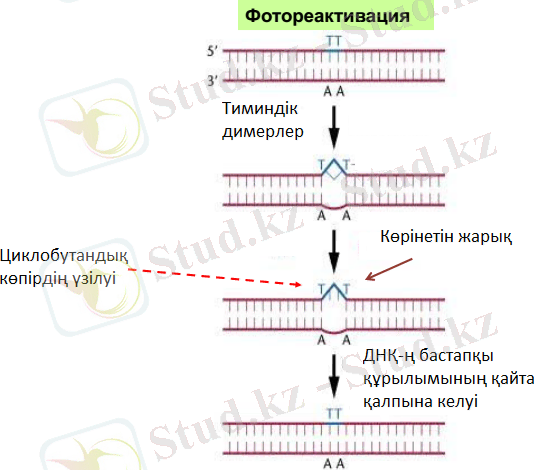
Фотореактивация құбылысы табиғатта кең таралған, тіпті микоплазма сияқты жабайы микроорганизмдерде де табылған. Фотореактивирлеуші ферменттер кейбір жоғары сатыдағы өсімдіктер мен жануарларда табылған, сондай-ақ барлық зерттелген бактериялардан табылған, оның ішінде УК-сәулелерге аса тұрақты Deinococcus radiodurans-ты ерекшелегенде, бұл бактериялар өлтіретіндерге қарағанда мөлшерді 1000 есе көтереді. Фотореактивацияға қабілеттігінің толық жоқтығынан Deinococcus radiodurans эксцизионды репарацияның күшті жүйесіне ие болады.
Бұзылған бөліктің алмастырушысымен байланысты репарациондық жағдайлар көрінетін жарықтың қатысуын талап етпейді және онда өзге ферменттерден басқа нуклеазаның екі типі маңызды рөлді ойнайды: экзо және эндонуклеазалар . Нуклеаза дегеніміз - қант-фосфатты тізбекті үзуге қабілетті ферменттер. Экзонуклеазалар тізбектің соңынан бастап ДНҚ-ның ыдырауын жүзеге асырады, ал эндонуклеазалар ДНҚ-ң бір тізбектік үзілістерін қалыптастыра отырып ішкі бөліктердегі тізбекке ұмтылады. ДНҚ-ның репаративтік синтезімен байланысы репарацияның түрлерінің әр алуандығының ішінен екі негізгісін белгілеуге болады: эксцизионды және пострепликативтік репарация .
ДНҚ-полимеразалардың экзонуклеазалық белсенділігі бойынша ДНҚ репарациясы. Бактериялық полимеразалардың көбісі (5’-3’ полимеразалық белсенділіктен басқалары) мүмкін қателіктердің коррекциясының арқасында 3’-5’ экзонуклеазалық белсенділікті иеленеді. Бұл коррекция екі кезеңде жүзеге асады: әуелі әр нуклеотидтің өсіп жатқан тізбек құрамына қосар алдында матрицаға сәйкестігін тексеру жүреді, кейін - нуклеотидті қосу жүреді. Дұрыс емес нуклеотидті қосу кезінде қос спираль деформацияланады. Бұл ДНҚ-полимеразаға көп жағдайда тізбектің дефектін тануға мүмкіндік береді. Егер қате салынған нуклеотид комплементарлы негіздермен сутекті байланысты қалыптастыруға қабілетті болмаса, полимераза репликация процесін керек нуклеотид оның орнына тұрмайынша тоқтата тұрады. E. coli-ден ДНҚ-полимераза III-ң ℇ-суббірлікті өзгертетін мутация - mutD генін анықтаған, нәтижесінде дұрыс салынбаған нуклеотидтердің генетикалық репарацияның бұзылысына, ақыр аяғында басқа гендерде мутацияның пайда болуына әкеледі.
2. 2 Эксцизионды репарация
Генетикалық репарацияның ДНҚ құрылымына “хирургиялық” араласуын еске түсіретін системасы болады: зақымдалған аумақтар ДНҚ тізбегінен кесіледі, осыдан «эксцизионды репарация» термині қалыптасады. Феноменнің өзі 1955 жылдан белгілі, алайда, эксцизионды репарацияның молекулярлық механизмі радиация әсеріне сезімтал мутантты бактериялар желісінде зерттеушілердің жұмыстарының нәтижесінде анағұрлым кеш - 1964 жылда ашылды. Генетикалық репарацияның бұл типі лигирлену мен кемшілікті құратын синтезге ілесетін ДНҚ-ң дұрыс емес немесе зақымдалған аумақты кесіп алуын қамтамасыз етеді. Бұл типке бірнеше арнайы механизмдер жатады, мысалы, гликолазалар тек түрлендірілген негіздерді ғана жояды, АР-эндонуклеазалар апуриндік сайттарды жояды және т. б. Көрініп тұрғандай, дәл эксцизиондық репарацияның жүйелері клеткадағы ДНҚ-ның зақымдалған бөлігінің үлкен аумағын қайта қалпына келтіреді[6] .
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz