Гендердің тіркесуі және кроссинговер: рекомбинация мөлшері, хромосомадағы орналасуы және механизмдері

Мазмұны
Кіріспе . . . 3
І. Негізгі бөлім
1. 1 . Тіркесе тұқымқуалау құбылысының ашылуы
1. 2. Тіркесу топтары. Кроссинговер . . .
1. 3. Алмасудың мөлшері және гендердің хромосомада тізбектеле орналасуы . . .
1. 4. Қос және көптік кроссинговер.
1. 5. Кроссинговердің цитологиялық дәлелдері
1. 6. Кроссинговердің механизмдері. Мейоздық кроссинговер …
1. 7. Тең емес кроссинговер. 1. 8. Кроссинговерге әсер етуші факторлар 1. 9. Кроссинговердің молекулалық механизмі
Қорытынды . . . 32
Қолданылған әдебиеттер тізімі . . . 33
Негізгі бөлім
1. 1 Тіркесе тұқымқуалау құбылысының ашылуы
Г. Мендельдің белгілердің тәуелсіз тұқымқуалау заңына сәйкес егер будандастырылатын дарабастардың екі немесе бірнеше жұп аллельдерінде айырмашылықтар болатын болса, онда әрбір жұп аллельдердің тұқымқуалауы бір-біріне тәуелсіз түрде жүреді. Әртүрлі гендердің осылай тәуелсіз таралуы мейоз кезінде аталық және аналық хромосомалардың әртүрлі жұптарының бір-біріне тәуелсіз түрде түрлі полюстарға ажырауына негізделген. Егер дигибрид АаВЬ болса, яғни олардағы екі жұп аллельдер әртүрлі хромосомаларда орналасса, онда А-а және В-в аллельдері жыныс клеткаларында бір-біріне тәуелсіз таралады және Ғ г -де фенотип бойынша ажырау 9 : 3 : 3 : 1 қатынасындай болады. Осындай қос рецессивті дигибридті талдау жасау үшін будандастырғанда алынатын ұрпақтартың фенотиптерінің арақатынасы 1 : 1 : 1 : 1 тең болады.
Енді гендердің екі аллельді жұбы Аа мен Вв екі әртүрлі хромосомаларда емес, хромосомалардың бір ұқсас ( гомологты ) жұбында ғана болады делік. Бірінші ұрпақтың гибридтерін өзара будандастырғанда
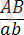
×
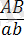
генотипі бойынша Ғ 2 -де болып ажырайтындығын байқаймыз. Егер Ғ 1 -дегі гибрид өсімдік талдаушы ретінде будандастырылса:
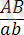
×
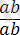
, онда Ғ 2 ұрпағында ажырау 1:1:1:1-ге емес 1 : 1 қатынасындай болып шығады ( ×2
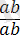
, ), яғни еркін, тәуелсіз комбинациялану жағдайында болатын төрт түрлі фенотиптік кластар байқалмайды. Бұдан шығатын қорытынды, бір жұп ұқсас хромосомаларда болатын гендер ұрпақтарға ажырамаған күйінде беріледі, яғни бірігіп тұқымқуалайды, себебі олар гаметогенез кезінде бір гаметада қалады, ал олардың Ғ а және Ғ 2 ұрпақтарының белгілері ата-аналарының формалары сияқты комбинацияда болады.
Белгілердің тәуелсіз комбинациялану заңының бұзылуын ең алгаш рет 1906 жылы У. Бетсон және Р. Пеннет анықтады. Хош иісті бұршақтың әр түрлерін будандастыра отырып олар дигибридті будандастыру кезінде екінші ұрпақта белгілердің ажырауынан күтпеген, өзгеше сипаттағы арақатынасты байқады. Қара қошқыл гүлді тозаңы үзынша келген ( RRLL) өсімдікті қызыл гүлді тозаңы дөңгелек (rrll) өсімдікпен будандастырғанда Ғ 1 -де қарақошқыл гүлді, тозаңы үзынша келген (RrLl) өсімдіктер алынған. Ғ 1 - дегі гибридтерді өздігінен будандастырғанда Ғ 2 -де мынандай ажыраулар болған:
Қарақошқыл гүлді, тозаңы ұзынша (R-L-) - 4831 (69, 5%),
Қарақошқыл гүлді, тозаңы дөңгелек ( R-ll) - 390 (5, 6%),
Қызыл гүлді, тозаңы ұзынша ( rr L-) - 393 (5, 6%),
Қарақошқыл гүлді, тозаңы үзынша (rr ll) -1138(19, 3%) . Бұл белгілерден ажырауға тиісті барлық төрт фенотиптік кластардың да алынғандығын көруге болады, бірақ та олар белгілердің тәуелсіз тұқымқуалауы кезіндегі дигибридті будандастыруға тән 9 : 3 : 3 : 1 қатынасындай емес. Кейінірек мұндай ажыраудың, егер Ғ 1 -дегі будандардан төрт түрлі гаметалардың түзілуі бірдей жиілікте жүрмесе ғана жүзеге асатындығы анықталды. Олай болса, зерттелген гендер аллельдерінің (RL және rl) ата-аналық үйлесімі негізінен бір ғана гаметанын үлесіне тиеді және бұл жағдайда олардың жаңа рекомбинантты үйлесімдері rL және Rl өте сирек кездеседі. Бұл құбылыс кейінірек гендердің тіркесуі деп аталынды. Әртүрлі белгілердің тұқымқуалауын көбірек зерттеген сайын гендердің тіркесу құбылысының заңдылықтарын түсінуге мүмкіншілік туды. Бұл құбылыстың мәнін түсіну мүмкіндігі тек тіркесудің материалдық негізі хромосома екендігін дәлелдейтіні Т. Морган зерттеулерінен кейін ғана туды. Хромосомалар клетканың редукциялық бөлінуі кезінде өз алдына жеке материалдық және функциональдық бірлік болып есептелінеді және гендер деп аталатын материалдық бөлшектерді алып жүруші болып саналады. Олай болса, бір хромосомада шоғырланған барлық гендер бір-бірімен тіркеседі.
1. 2 Тіркесу топтары. Кроссинговер
Гендер тіркесу құбылысы ашылғаннан кейін Т. Морган мен оның қызметтестері бір жұп гомологты хромосомада орналасқан гендердің тұтасымен тұқымқуалап тіркесу топтарын құрайтындығын анықтады. Белгілі бір топқа жататын гендерді будандастыру кезінде бір-бірімен тіркесетіндігі байқалды, ал әртүрлі топтарға жататын гендер бір-біріне тәуелсіз тұқым қуалайды. Жүргізілген зертгеулердің нәтижесі гендердің тіркесу топтарының саны хромосомалар жұптарының санына сәйкес келетіндігін көрсетті. Мысалы, дрозофила шыбынында 4 жұп хромосома және 4 тіркесу тобы, арпа мен бұршақта 7 жұп хромосома және 7 жұп тіркесу тобы, жүгеріде 10 жұп хромосома және 10 жұп тіркесу тобы, қызанақта (томат) 12 жұп хромосома және 12 жұп тіркесу тобы бар. Кейіннен бұлардан басқа да көптеген өсімдіктер мен жануарларда тіркесу топтары зерттелді. Адамның 23 жұп хромосомаларына сәйкес келетін 23 тіркесу топтарының болатындығы анықталды. Осы күнге дейін тіркесу топтарының саны хромосома жұптарының санынан артық болып келетін бірде-бір организм табылмаған.
Әрбір жеке организм әртүрлі гендердің көптеген аллельдері бойынша бір-бірінен айырмашылықтары бар аталық және аналық гаметалардың қосылуы нәтижесінде пайда болады. Жоғарыда аталып көрсетілгендей екі тіркескен гендер бойынша айырмашылықтары бар организмдерді будандастырғанда: :
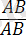
×
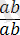
, дигетерозиготалы форма
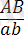
пайда болады.
Толық тіркесу жағдайында дигетерозигота тек екі түрлі ғана гаметалар түзеді: АВ және ав. Талдаушы будандастыру кезінде зиготалардың екі
класы пайда болады:
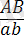
және
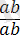
, олардың әрқайсысы 1 : 1 тең болады.
Екі кластың да дарабастары өздерінің ата-аналарының белгілерін қайталайды. Мұндай көрініс талдаушы будандастыру кезінде дигибридті ажырау емес моногибридті ажырауды еске түсіреді.
Дегенмен толық тіркесу құбылысымен қатар заңды түрде толымсыз тіркесу құбылысы да болып отырады. Толымсыз тіркесу құбылысы жағдайында генотипі
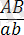
гетерозиготалы дарабастарды рецессивті
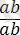
формалармен будандастырғанда олардың ұрпағында жаңа белгілері бар дарабастар пайда болады:
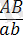
және
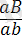
. Мұндай жаңа фенотиптер рекомбинанттар деп аталады. Ажырау барысында зиготалардың жаңа кластарының пайда болуы екі ген бойынша гетерозиготалы формалардың гаметогенезі процесінде тек қана АВ және ав емес, сонымен қатар Ав және аВ гаметалары да түзілетіндігін көрсетеді. Олай болса Ғ 1 буданына бір ғана хромосомамен келген гендер, одан гамета түзілу кезінде қандай да бір жолмен ажырап кетеді. Гендердің жаңа үйлесімі бар мұндай гаметалары қалай пайда болды? Демек, олар тек гомологты хромосомалардың арасында бір-бірімен бөлімдерін алмастыру, яғни кроссинговер кезінде пайда болуы мүмкін. Сонымен кроссинговер ұқсас (гомологты) хромосомаларда болатын гендердің жаңа үйлесімін қамтамасыз етеді. Кроссинговер құбылысы да тіркесу сияқты барлық жануарлар мен өсімдіктерге және микроорганизмдерге тән құбылыс.
Кроссинговер тек гендер гетерозиготалы (
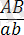
) болғанда ғана байқалады.
Гендердің гомозиготалы
.
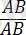
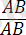 және
және
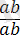
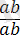 қалпында хромосомалардың айқасуын көре алмаймыз, себебі, ұқсас гомологты бөліктердің алмасуы гаметаларда және ұрпақта гендердің жаңа комбинацияларын бермейді.
қалпында хромосомалардың айқасуын көре алмаймыз, себебі, ұқсас гомологты бөліктердің алмасуы гаметаларда және ұрпақта гендердің жаңа комбинацияларын бермейді.
Хромосомалардың айқасуы туралы, пайда болған рекомбинанттардың жиілігіне генетикалық талдау жасау негізінде ғана, яғни мейоздағы хромосомаларға цитологиялық зертгеулер және гендердің жаңа үйлесімі бар зиготаларға талдаулар жүргізу арқылы ғана пікір айтуға болады. Тіркесу және айқасуға мысал ретінде К. Гетчинсонның жүгерімен жүргізген тәжірибесін қарастырайық. Тұқымы тегіс, боялған өсімдікпен (ССSS) тұқымы бұдырлы боялмаған өсімдік (ccss) будандастырылған. Тұқымының тегістігі (С) мен бояуы (S) доминантты белгілер, сондықтан бірінші ұрпақта барлығы да боялған тегіс тұқымды (СсSs) болады.
Осындай тұқымдардан өсіп шыққан өсімдіктер рецессивті (ақ түсті бұдырлы) дарабастармен будандастырылған:
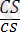
×
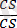
. Сонда алынған ұрпақтың төмендегідей сандық арақатынаста төрт түрлі тұқымдарының болатындығы байқалған:Басқаша айтқанда, бастапқы екі түрдің тұқымдары олардың барлық сандық мөлшерінің 96, 4%-ін, ал тұқымдарының қалған екі тобы (кроссинговердің нәтижесінде пайда болған) 3, 6%-ін құрайды. Бұл тәжірибенің нәтижесі, шындығында гендер тіркесуінің бар екендігін, бірақ кейде кроссинговердің салдарынан оның аздаған мөлшерде өзгеретіндігін, яғни мейоз кезінде ұқсас гомологты хромосомалардың арасындағы біркелкі бөлімдерінің алмасатындығын көрсетті. Ұқсас гомологты хромосомалардың бірдей бөлімдерінде орналасатын гендер олардың біреуінен екіншісіне ауысып отырады.
Боялған тегіс
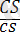
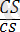 )
)
-4032
Ақ түсті бүдырлы
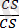
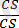 )
)
-4035
Боялған бүдырлы
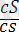
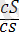 )
)
- 152
Ақ түсті тегіс
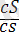
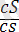 )
)
- 149
Гендер тіркесуі мен кроссинговерге мысал ретінде Т. Морганның тәжірибесін де келтіруге болады. Дрозофила шыбынының денесінің қара түсін бақылайтын ген bаск деп аталады және ол b әрпімен белгіленеді Оның доминантты аллелі - b + Рецессивті vg (vеstgial) гені гомозиготалы жағдайда қанатының жетілмегендігіне (тұқыл қанат) әкеп соғады. Оның доминантты аллелі - Vg + қанаттарының дұрыс жетілуін бақылайды. Денесі қара қанаты жетілмеген шыбынды, денесі сұр қанаттары қалыпты шыбындармен будандастырғанда :
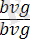
×
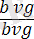
Ғ 1 - де осы гендер бойынша дигетерозиготалы дарабастар алынған: :
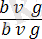
. Кейіннен екі түрлі талдаушы будандастыру жүргізілген: біреуінде дигетерозигота аталық, екіншісінде аналық болған. Егер гибрид аталықтар екі гендер бойынша да рецессивті аналықтармен будандастырылса, яғни талдаушы будандастыру
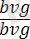
×
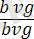
жүргізілсе, онда ұрпағында жынысына ешқандай байланысы жоқ екі түрлі шыбындар алынады, яғни шыбындардың 50% қара денелі қанаттары жетілмеген (
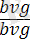
), ал қалған 50% екі белгі бойынша да қалыпты болады. Мұндай ажырау осы дигибридтің ата-анасынан берілген аллельдердің негізінде тек екі сортты ғана гамета түзетіндігін ( b + vg + және bvg ) көрсетеді. Тәжірибе жүзінде дрозофила шыбынының аталықтарының аутосомаларында және жыныстық хромосомаларында да қалыпты жағдайда гомологты хромосомалардың бөлім алмастырмайтындығы анықталды. Сондықтан талдаушы будандастырудың бұл түрінде 100% жағдайда зерттелген гендер бойынша тек ата-анасына сай келетін гаметалар түзілген. Бұдан біз бір жұп гомологты аутосомаларда болатын гендердің толық тіркесетіндігін көреміз. Дигетерозиготалы аналықты екі рецессивті гендер бойынша гомозиготалы талдаушы (анализатор) аталықпен реципрокты будандастырғанда ұрпақтың болуға тиісті төрт түрі де көрініс береді. Белгілердің ата-аналық комбинацияларынан басқа, ұрпағында шыбындардың жаңа түрлері - қара денелі қалыпты қанатты ( bvg) + және сұр денелі қанаттары жетілмеген (b + vg ) пайда болады. Олай болса, бұл будандастыруда гендер тіркесуі бұзылады. Гомологты хромосомалардағы гендер хромосомалардың айқасуына байланысты орын алмастырады. Кроссинговерге ұшыраған хромосомалары бар гаметалар кроссоверлі, ал кроссинговерге ұшырамағандар кроссоверлі емес деп аталады. Ажырауға талдау жасау кезінде, екінші будандастыруда кроссинговер жағдайында әртүрлі класты дарабастардың белгілі бір сандық ара қатынаста болуы озіне көңіл аудартады. Кроссоверлі емес гаметалардан пайда болған бастапқы атааналық белгілердің екі комбинациясы талдаушы будандастырудың нәтижесінде алынған ұрпақта сан жағынан алғанда тең (41, 5%) болып шығады. Сонда ата-анаға сай келетін кроссоверлі емес гендердің bvg және b + vg + жиынтығы 83% құрайды. Ал жаңа комбинациялар, яғни bvg + және b + vg рекомбинациялары 17% құрайды.
Басқа тәжірибеде ата-аналар ретінде дәл сондай белгілері бар, бірақ басқа үйлесімділіктегі шыбындар алынған: қара денелі қалыпты қанаты
(
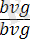
) шыбынды, сұр денелі қанаты жетілмеген (
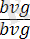
) шыбынмен
будандастырған. Содан кейін қайтадан екі түрлі талдаушы будандастыру жүргізілген. Бірінші будандастыруда Ғ 1 -дің аталықтары, екіншісінде Ғ 1 -дің аналықтары пайдаланылды. Екеуінде де олар қос гомозиготалы
рецессивпен (
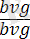
) будандастырылған. Сонда қайтадан, талдаушы
будандастыру кезінде ата-аналық және рекомбинантгық белгілердің арақатынасы бұрынғыдай болып шыққан. Егер Ғ 1 -дегі аталықтар алынса, онда белгілердің тек қана ата-аналық комбинациясы, ал егер Ғ 1 -дегі аналықтарды алса, онда 83% ата- аналық және 17% рекомбинациялық белгілер байқалады, яғни белгілердің үйлесімі алғашқы тәжірибедегідей ара қатынаста болады.
1. 3 Алмасудың мөлшері және гендердің хромосомада тізбектеле орналасуы
Рекомбинацияға ұшыраған дарабастарды және олардың жалпы санын ескере отырып, кроссинговер (айқасу) жиілігін мына формуламен есептеп шығаруға болады:
кроссинговер жиілігі ('%) =

 х 100
х 100
Мұндағы Х-рекомбинацияға ұшыраған дарабастар саны; У- дарабастардың жалпы саны. Мысалы, Ғ 1 ұрпағында алынған шыбындарды гомозиготалы қос рецессивтілермен қайыра будандастырғанда мынандай нәтижелер алынған:
Рекомбинация жиілігі:
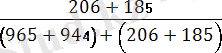
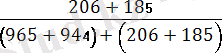 ×100=
×100=
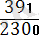
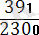 ×= 17%
×= 17%
Бұл мән гаметалар түзілуі кезінде пайда болатын рекомбинация санына сәйкес келеді. Айқасудың мөлшері хромосомадағы гендер тіркесу күшін көрсетеді: неғүрлым айқасу мөлшері жоғары болса, тірк күші соғүрлым аз болады. Т. Морган кроссинговердің жиі хромосомадағы гендердің
орналасу сипатын көрсетеді деп тұжырымда яғни тіркес гендер бір-бірінен неғүрлым алшақ орналасса, олар ; арасында кроссинговердің болу мүмкіндігі соғүрлым көбе рекомбинантгардың жиілігі арта түседі.
Біз дрозофила шыбынының денесінің қара түсі мен қанаты жетілмегендігін анықтайтын гендердің рекомбинациялану жиілігі 17% көрсеткенде, бұл шама хромосомадағы осы гендердің арасынд қашықтықты сипаттайтын болады. Мұндай жағдай жұп хромосомаларында болатын айқаста да тән, яғни анықталған 3 рекомбинация гомологты хромосомалардың екі бөлімшесінің арасында болатын жиілікті көрсетеді.
Кроссинговер құбылысы, гендер хромосомада бір сызықтың бойы белгілі бір ретпен орналасады деген тұжырымға әкеледі. Гендер тек сызық бойында орналасса ғана рекомбинанттардың пайызы хромосомад гендердің салыстырмалы түрдегі ара-қашықтығын көрсете алады. геннің арасындағы кроссинговердің жиілігіне қарай, хромосомадағы гендердің бір-бірінен ара-қашықтығы туралы пікір айтуға болады.
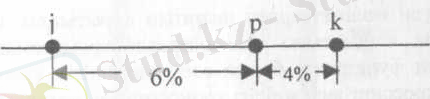
немесе
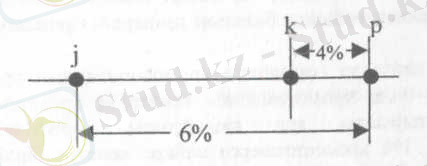
№1-сурет. Гендердің хромосомада орналасу ретін анықтайтын әдістері сызбажобасы.
Бұл ұғымның дұрыстығын нақты мысалмен тексеруге болады. Айталық, дрозофилада
j - жоғары қарай қайқайған қанат, р - көздің қара қошқыл түсі
және к - көздің қызыл түсі сияқты үш түрлі мутациялар кездеседі. Зерттеудің нәтижесінде бұл мутациялардың бір хромосомада шоғырланған гендерге байланысты екендігі және осы гендердің арасында тіркесудің болатындығы анықталған. Жүргізілген тәжірибелер j және р гендерінің арасында 6% кроссинговер, ал р мен к гендерінің арасында 4% кроссинговер болатындығын көрсетті. Бірақ бұл мәліметгер j, р және к гендерінің қандай ретпен орналасатындығын айтуға мүмкіндік бермейді (№1-сурет) .
Бұл мәселені шешу үшін j мен к гендерінің арасындағы кроссинговердің процентін анықтау керек. Бұл екі ген арасындағы кроссинговер 10%-ке тең, яғни ол сан бұрыңғы алынған (j мен р және р мен к) гендердің арасындағы кроссинговердің қосындысына тең. Ол дегеніміз суретте к гені р генінің оң жағына қарай орналасуы дұрыс екендігін көрсетеді
Осы берілген мәліметтерден шығатын корытынды, айқасудың пайызы гендердің ара қашықтығы мен олардың хромосомада сызық бойына орналасуының функциясы болып есептелетіндігін көрсетеді, яғни гендер арасындағы кроссинговер жиілігі хромосомадағы оларды бөліп тұратын ара қашықтыққа тура пропорционалды және осы мәліметтердің негізінде хромосомалардың генетикалъщ картасын жасауға болады. Генетикалық карта деп бір тіркесу тобында болатын гендердің орналасу сызбажобасын айтады.
Генетикалық карталар гомологты хромосомалардың әр жұбына жеке- жеке жасалады, онда хромосомадағы гендердің орналасу реті мен бір- бірінен салыстырмалы ара қашықтығы көрсетіледі. Бұл ара қашықтықтарды 1% кроссинговерге сәйкес келетін бірліктерде көрсету мақұлданған. Бұл бірліктерді алғаш рет хромосомалардың картасын құрған Т. Морганның құрметіне морганида деп атайды. Генетикалық картаны құруда әдетге, әр
объектіге тән гендерді белгілеудің белгілі бір жүйесі қолданылады. Бірақ карта құрудың жалпы үлгісі бірдей. Онда міндетті түрде тіркесу топтары, гендердің толық немесе қысқартылған атаулары,
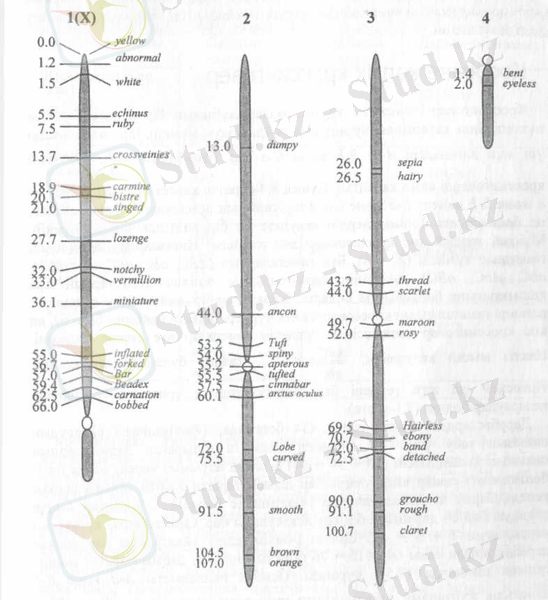
№2-сурет. Дрозофила хромосомаларының генетикалық картасы. 1-иіі хромосома Х-хромосомасы
хромосоманың бір жақ шетінен басталатын морганидамен белгіленетін ара қашықтық көрсетіледі. (№2-сурет)
Генетикалық тұрғыда біршама жақсы зерттелген организмдердің, мысалы, дрозофила шыбыны, жүгері, бұршақ, нейроспоралардың барлық хромосомаларының карталары жасалған және оларды бүкіл гендердің орналасқан орындары көрсетілген. Қазіргі кезде адамның да барлық хромосомаларының генетикалық картасы жасалынып, гендердің орналасу реті анықталған.
1. 4 Қос және көптік кроссинговер
Кроссинговер гомологты хромосомалар жұбының бір немесе бірнеше нүктелерінде қатарынан бір мезгілде пайда болуы мүмкін. Бір хромосомада үш жүп аллельдері А-а, В-в және С-с бар үшгибридте болатын
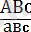
болатын кроссинговерді алып қарайтын болсақ 8. 4-суретте көрсетілгендей айқасу А-а ; және В-в немесе В-в және С-с локустарының арасындағы бөлімшелерде, болмаса екі бөлімшелердің екеуінде де бір мезгілде болуы мүмкін. Ондай жағдай қос кроссинговер деп аталады. Нәтижесінде сегіз түрлі гаметалар түзіледі (2 3 = 8) . Бұл гаметалардың (АВС, авс, аВС, АВс, Авс, АВс, аВС) түзілуінің салыстырмалы жиілігі локустардың ара қашықтығына байланысты болады. Бірақта, әрбір жеке нақты жағдайды алғанда гаметалардың кроссоверлі емес түрлері біршама көбірек болады, ал кроссинговер нәтижесінде түзілетін гаметалар өте сирек кездеседі.
Нақты мысал келтірейік:
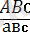
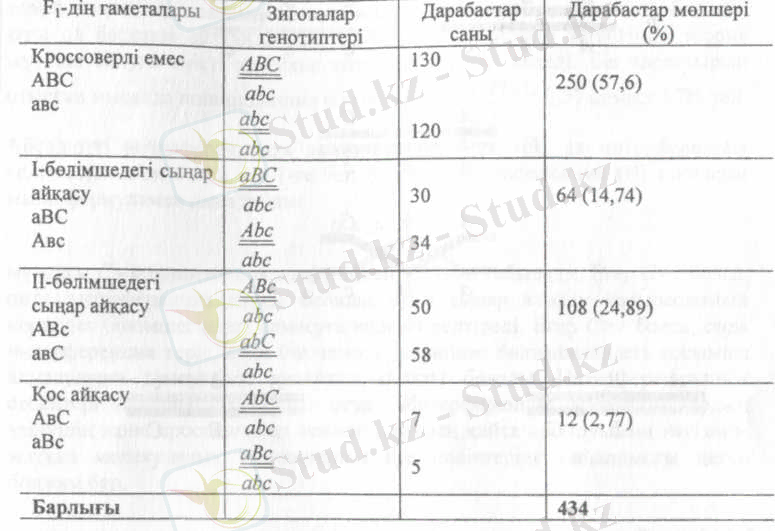
×
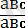
талдау будандастыру кезінде тірк е скен үш жұп гендері бар үшгибридтердің ұрпағында мынандай ажыраулар болған (№1-кесте) .
Дарабастардың жалпы саны 434 болғанда, І-бөлімшедегі айқасудың ұзын табу үшін біз алдымен айқасуға ұшыраған дарабастардың санының қосындысын (30 + 34 = 64) есептеп алуымыз керек, сонда ол I- бөлімшедегі сыңар айқасулары бар дарабастардың жалпы санына сәйкес келеді. Бірақ қос айқасулар да І-бөлімшеде болады. Сондықтан сыңар айқасуы бар 64 дарабасқа біз қос айқасулары бар дарабастарды қосуымыз к, яғни 7 + 5 = 12. Сөйтіп І-бөлімшедегі айқасулары бар жалпы бастардың саны 64 + 12 = 76 болады. Бұл сан дарабастардың жалпы санына шаққанда 17, 5% құрайды. Осыған байланысты А-а және В-в
алельдік жұптардың локустарының арасындағы қашықтық:
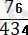
× 17, 5% морганида болып шығады.
Дәл осындай жолмен ІІ-бөлімшедегі айқасулары бар дарабастардың жалпы санын да анықтауға болады. (50 + 58 + 7 + 5 =120) . Бұл санды дарбастардың жалпы санына шағып пайыз есебімен алғанда II- бөлмішедегі айқасудың жиілігі 27, 6%-ға тең болады. Сонымен, осындай жолмен гендердің арасындағы салыстырмалы қашықтықты анықтаймыз: А мен В гендерінің ара қашықтығы 17, 5, ал В мен С - 27, 6 морганида болады. Мұнда қос айқаспалы 12 дарабас І-бөлімшенің және ІІ-бөлімшенің де айқасу жиілігін есептеген кезде еске алынғандығын ескеру керек.
№1-кесте талдаушы будандастыруда тіркескен үш жұп гендері бар үшгетерозиготаның ұрпағындағы ажырау.
Кроссинговер жиілігі гендер арасындағы қашықтықтың функциясы болғандықтан А және С гендерінің ара қашықтығы А-В және В-С гендері ара қашықтықтарының қосындысына тең болуы тиіс, яғни 17, 5+27, 6=45, 1% болады. А мен С гендерінің арасындағы сыңар кроссоверлердің жалпы саны 172 (64 + 108) дарабастарды немесе 39, 6% құрайды. Айқасудың бұл жиілігін I және ІІ-бөлімшелердегі бұрынғы есептелген айқасулар жиынтығымен салыстыруға болады. Олардың мәні 17, 5% және 27, 6%-ке тең болған немесе қосындысы 45, 1%, яғни А мен С локустарының арасындағы кроссинговердің жиілігін анықтау арқылы алынған мөлшер 45, 1-39, 6=5, 5% бірлікке көп екендігін көреміз.
Есептеудегі болатын келеңсіздіктерден алшақ орналасқан гендердің арасындағы қос айқасудың ара қашықтығын дәлірек анықтауға қиындық туатындығын байқалады. Егер А мен С гендерінің ара қашықтығы үшінші В генімен белгіленбесе, қос кроссинговер байқалмай да қалуы мүмкін. А- дан С-ға дейінгі қашықтық мынандай жолмен анықталады: сыңар кроссоверлі кластар пайыздарының қосындысына (39, 6%) қос
кроссоверлердің екі еселенген пайызын қосу керек (
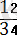
х 100 = 2, 77 х 2 =
5, 5%) . Сонда қорыта келгенде 39, 6 + 5, 5 = 45, 1% болады немесе ол екі бөлімшелердегі яғни А- дан В-ға және В-дан С-ға дейінгі кроссинговердің пайыздарын қосқанда алынатын жиынтыққа тең болып шығады. Екі геннің арасындағы рекомбинацияның пайызы неғүрлым щың ара қашықтығы кеміген сайын нақтылана түседі, себебі қашықтық азайған жағдайда қос алмасулардың болу мүмкіндігі де кемиді.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz