Дезоксирибонуклеин қышқылының құрылымы, қасиеттері және репликациясының молекулалық негіздері

Кіріспе
Тірі организмдер табиғи қасиеттері мен белгілерін ұрпақтан ұрпаққа беріп отыратынын адам баласы ертеден байқаған.
ХVIII ғасырдың барысында неміс зоологы А. Вейсман жыныс клеткаларында болатын ерекше заттар-тұқым қуалаушылықтың негізі болуы керек деп, оның құпиясын клеткалардағы молекулалардан іздеу туралы дұрыс ұсыныс жасаған еді.
Генетика тұқым қуалаушылықтың материалдық негізгі ең алдымен хромосомалар болатынын сенімді түрде көрсетіп берді. Құрамында гендері бар хромосомалар өз көшірмесін қалдыратын қатар түзеді. Тіршілікке тән үздіксіз көбею, өсіп-өну қасиет осы хромосомаларға байланысты.
Хромосомалардың өз көшірмесін қалдыратыны жөніндегі үлгіні 1928 жылы Н. К. Кольцов ұсынған. Ол «Om-»-әрбір молекула молекуладан деген жорамал айтқан. Бұл постулат бойынша клеткадағы макромолкулалар:белоктар және нуклеин қышқылдары матрицалық (көшірмелік) принциппен көбеюге тиіс. Хромосоманың құрлысы күрделі. Оның құрамына белоктар (гистон тәріздес және гистон тәрізді емес ), лицидтер, екі валентті металдар қатиондары т. с. с. кіреді. 1940 жылдың басына дейін хромосомалардың генетикалық қызметін көп зерттеушілер тек қана белокпен байланысты деп есептеген. Н. К. Кольцовтың айтуынша, «ДНҚ сияқты « қарапайым молекула» соншаман күрделі қызмет атқарады деп мойындау өте қиын еді. Бірақ кейініректе барлық өсімдіктерде, жануарларда, микрооганизмдерде, көпшілік вирустарда генетикалық материал-ДНҚ екені анықталды[1] .
Бүкіл организмдерде болатын тұқым қуалаушылық қасиет нуклеин қышқылдарының қызметтеріне байланысты. Олардың барлығының дерлік құрамынан ДНҚ табылған, тек кейбір вирустарда ғана оның орнына РНҚ болады. Нуклеин қышқылдары- күрделі биологиялық полимер. Олардың мономерлері нуклеотиттер болып табылады. Әр нуклеотид үш копманенттен тұрады: азотты негіздерден, пентоза қанттынан және фосфор қышқылынан. Азотты негіздердің бес түрі бар. Соның бірі урацил, ол тек РНҚ-ның құрамында кездеседі. Келесі тимин, бұл тек ДНҚ-да болады. Ал қалған үш азотты негіздер:цитозин, аденин және гуанин -ның да, РНҚ-ның да құрамына енеді. Екі циклді негіздер - аденин мен гуанин-пурин, ал моноциклді негіздер - цитозин, тимин және урацил пиримидин туындыларына жатады.
РНҚ-ның құрамына енетін пентоза қантты- рибоза ал ДНҚ-ның құрамындағы дезоксирибоза болып табылады. Осыған байланысты олар рибонуклеин және дезоксирибозануклеин қышқылдары деп аталады. Нуклеиотидтің қантпен азотты негізден тұратын бөлігін нуклеозид деп атайды. Соған байланысты нуклеотидтерді кейде деп атайды[2] .
Негізгі бөлім
1. 1 Нукеин қышқылдарының құрлымы және биологиялық ролі
Нуклеин қышқылдарының проблемасымен 1868 жылы швейцарлық физик Ф. Мишер шұғылданған болатын. Ол ірің клеткаларынан алынған ядро бөлшегін зерттеп, құрамына көміртегі, азот және фосфор кіретін бұрын белгісіз органикалық қосылыс түрін ашты. Ашқан қосылысын ядродан бөліп алғандықтан нуклеин деп атады (латынша-«нуклеус»-ядро) .
Ф. Мишердің ашқан қосылысы басқа да ғылымдардың назарынан тыс қалған жоқ. 1874 жылы Пиккард барлық спермасынан бөлініп алынған нуклеин құрамынан жаңа азоттық негізді ашты. Ол-құрамында екі азот атомы бар гетероциклдік қосылыстардың ішіндегі пуриндер тобына жататын гуанин еді.
Түсінікті болуы үшін гетероциклдің не екеніне және оны не себептен негіз деп аталатынына тоқтала кеткен жөн. Циклді қосылыс деп қаңқасы сақина түрінде тұйықталған көмірсутектерін айтады. Ал оның құрамына көмірсутектерінен басқа да атом түрлері кірсе, оны гетероциклді дейді (гетеро-әр түрлі) . Ал пуриндерді, пиримидиндерді негіз дейтіні-олардың құрамындағы азот қышқылды ортада өзңне протнды қосып алып оң зарядталады (цикдағы көмірсутектері ешқашан зарядталмайды) . 1880 жылы неміс химигі Г. Фишер әлі нукленнің құрамына тек пуриндік емес, сонымен қатар пиримидиндік гетероциклдер кіретінінанықтады. Пуриндер алты және бес атомды гетероциклдерден біріккен болса, пиримидиндер тек алты атомнан тұратын гетероциклді қосылыс. Бірнеше жылдан кейін Ф. Мишердің шәкірттері Коссель мен Асколи тағы төрт түрлі негіздерді-аденин, тимин, цитозин және урацилді ашты. Нуклеиннің ерітіндісі қышқылдық қасиет көрсететін болғандықтан Ф. Альтман 1889 жылы оған нуклеин қышқылы деген ат берді. Осы ғасырдың басында жүргізген зерттеулерінің нәтижесінде П. Левен нуклеин қышқылының құрамына азоттық негіз бен фосфордан басқа шағын углеводтардың ерекше тобы кіретінін анықтады (сахар) . Нуклеин қышқылының құрамына кіретін углевод бес көміртегінен тұрады. Левен нуклеин қышқылының бір тобына кіретін углеводтағы оттегінің саны бір атомда кем екенін анықтады. Оттегі атомдары толықтарын риб о за деп, бір оттегі атомы кемді-дезоксирибоза («дезокси»-оттексіз) деп атады. Сонымен, оттегінің бір атомының санындағы айырмашылық негізінде нуклеин қышқылдарының «кірпіштері»болып табылатынын анықтады. Нуклеин қышқылдарын құрайтын бұл «кірпіштерді» нуклеотид деп атады[3] .
1928 жылы Ф. Гриффит өкпе ауыруларын қоздыратын пневмококк бактериясының өсуін зерттегенде, олардың колониялары қоректік ортада агарда өзінің бейнесін өзгертетінін байқады. Алғашқыда тегіс, әрі жылтыр келген пневмококтардың «S» түрі (ағылшынша Smootn-тегіс), кейін бұдырмақтанып «R» (rugn-бұдырмақтау) түріне айналатынын байқалды. Бұл ауысу (S-R) диссциация деп аталды. Келесі жаңалық бірнеше жылдан кейін ашылды-пневкоктарды кері бағытта R-күйден S-күйіне өзгертуге мүмкіндік туды. Диссоциацияға қарағанда кері бағытта өтетін бұл өзгеріс трансформация деп аталды.
1944 жылы Американдық микробиологтар К. Т. Эйвери, К. М. Маклеод және М. Маккарти трансформацияны туғызатын пневмококтарда болатын дезоксирибонукленин қышқылы деп дәлелдеді.
Міне, осы 1944 жыл молекулярлық биологияның ашылған жылы деп есептенінеді.
50-жылдардың басында Э. Чаргафф өте маңызды жаңалық ашты. Белоктар тәрізді нуклеин қышқылдарының да әр түрге тән екенін анықтау еді.
Э. Чаргафф өте таза ДНҚ молекуласын бөліп алып, мұқият түрде химиялық талдау жасап, кез келген организмнен бөлініп алынған ДНҚ-ның құрамындағы адениннің мөлшері тиминдікімен (А=Т), ал гуаниннің мөлшері цитозиндікімен (Г=Ц) бірдей болатынын анықтады.
(А+Г)
Г+Ц
Осы ережеге сәйкес әр түрі организмдердің нуклеотидтік құрамы тек қана олардың арқатынасының өзгеруіне, А+Т мөлшеріне байланысты.
1952 жылы Р. Франклин және М. Уилкинс дНҚ-ның жоғарғы сапалы рентгенограммасын түсірді.
Осы ренгенструктуралық талдаудың және ДНҚ-ның химиялы құрамын біле отырып, 1953 жылы Д. Уотсон және Ф. Крик оның молекулалық моделін құрастырды. Бұл биология тарихындағы ең үлкен жаңалықтың бірі болып табылады. Ол аденин мен тиминнің, гуанин мен цитозиннің бір-бірімен байланысы. Мұнда модель Чаргаффтың заңдылығына сай келеді.
Х. Резерфордтың атомның мқұрлысын ашуы адамзатқа шексіз энергия кһзін берсе, Уотсон-Крик және Уилкинстердің ДНҚ-ның құрлысын ашуы организмге жаңа қасиет бере алатын ген инженериясының әдісін ашты[4] .
1. 1. 1 Дезоксирибонуклеин қышқылы - ДНҚ
Организмдердің басым көпшілігінде (құрамына РНҚ кіретін вирустерден басқа) тұқым қуудың материалдық негізі болып табылады. Ол адамның және барлық басқа организмнің клеткаларында болады. Сонымен бірге ДнҚ кейбір бақтериофагтар мен вирустарда да болады. Организмдегі барлық клетка ядроларында ДНҚ саны әрдайым бірдей болады.
ДНҚ-ұзын макромолекула, оның негізгі құрылыс мүшелері болып табылады. Нуклеотид үш құрылымды элементтен: органикалық қосылыстан, углеводтан және фосфор қышқылынан құралған. Днқ-да дезоксирибоза деп аталатын углевод пен фосфор қышқылы барлық нуклеотидтерде бірдей; нуклеотидтерде органикалық қосылыстар бірнешеу: аденин (А), тимин (Т), гуанин (г) және цитозин (Ц) . ДНҚ молекуласы ширатылған қос тізбекті спираль (латынша «спиралис»-шиыршық сым) тәрізді иірілген. Әр тізбектің нуклеотидтерінің бір-бірімен кезектесіп жалғасқан углевод пен фосфаттардың қатары осы спиральдың қаңқасы болып табылады. Шиыршықтың ішкі кеңістігінде бір тізбектің азоттық негіздері екінші тізбекке сйәкес негіздермен сутегілік байланыстар құрап, бір-бірімен берік ұстасып сутегілік байланыстар құрап, бір-бірімен берік ұстасп тұрады. Азоттық негіздердің химиялы құрлыстарының ерекшеліктері бір тізбектің аденні (А) екі сутегілік байланыс нәтижесінде екінші тізбектінң тек тиминімен (Т( ғана, ал гуанин (Г) үш байланыс арқылы тек тиминімен (Т), ал гуанин (Г) үш байланыс арқылы тек цитозинмен (Ц) ғана біріге алатынын көрсетеді. Яғни, бір тізбектінң бойындағы азоттық негіздердің қандай кезекпен орналасқаны белгілі болса, онда ол тізбекпен бірігіп тұрған екінші тізбектің бойындағы азоттық негіздердің орналасу тәртібін де өте оңай анықтауға болады. Осындай бірімен бірі сәйкес келетін тізбектерді бір-біріне комплементарлы ((комплементарлы-сәйкес, толықтырушы) деп атайды. ДНҚ-ның шиыршық қаңқасы үнемі қайталанып отыратын углевод-фосфат тобы болса, оның ішкі кеңістігінде біріккен азоттық негіздердің жұбы бірінен кейін бірі спиральдың бойында әр түрлі кезекпен орналасады . ДНҚ тізбектерінің бағыты бір-біріне қарама-қарсы. Спиральдің бір ұшында бір тізбек углеводтың 3-көміртегінің гидроксил тобымен бірте, екінші тізбек 5-көміртегімен байланысқан фосфатпен аяқталады. Ал спиральдің ол тізбектерінің екінші ұштары керісінше аяқталған, сондықтан ол тізбектердің бағытын былай өрнектейді:3-5және 5-3, яғни олардың бағыты қарама-қарсы деген сөз. ДНҚ молекуласы шиыршығының формасы (сырт пішіні) сағат тілінің бағыты бойымен оңға қарай ширатылған, ол бұралған баспалдаққа ұқсайды. Екі жіпшенің арасындағы жұптасқан А-Т және Г-Ц топтары ол баспалдақтың басқыштары секілді. Шынында да ДНҚ молекуласында ол «басқыштар»бір-біріне параллель, ал спираоьдың ұзына бойы осіне перпендикуляр орналасқан. ДНҚ молекуласы неліктен спираль формалы? Бір тізбектегі азоттық негіздердің барлығы дерлік екінші тізбектіңбарлық негіздерімен толық жақындасып байланеысу үшін ол тізбектер бір-біріне оралып спираль болып айналуы шарт екен. Сонымен қатар спираль ішіндегі параллель орналасқан жұп азоттық негіздердің арасында гидрофобтық (грекше «гидро»-су, «фобос»-қорқу ), және дисперсиялық (латынша «дисперсус»-шашылу, ыдырау) күштердің әсерімен қосымша байланысты- стэкинг байланысы деп атайды. Мұндай байланыс пуринді азоттық негіздердің арасында күштірек. ДНҚ-молекуласын спираль күйінде ұстап тұратын осы байланыстар[5] .
Ал енді ДНҚ-ның мөлшері туралы не айтуға болады? ДНҚ-ның спиралінің диаметрі 20 ангстрем. Бір-бірімен параллель жатқан көрші жұп негіздердің ара қашықтығы 3, 4 ангстрем. Спиральдың бір айналымына жұптасқан 10 азоттыөқ негіздер кіреді, яғни спиральдың бір айналымы ұзын ДНҚ молекуласының бойындағы 34 ангстрем бөлікті алып жатыр. Ал ДНҚ-ның ұзындығы қандай? Ол ДНҚ-ның қандай организмнен алынғанынанбайланысты: қарапайым вирустың ДНҚ-сы бірнеше мың жұп негізден тұрса, бактериялардікі бірнеше миллион, ал жануарлардікі көптеген миллиард негіздерден тұрады. Адамның бір клеткасындағы барлық ДНҚ молекулаларын созса, ұзындығы 2 метірге жетеді, яғни диаметірі 20 ангстрем, ұзындығы 2 метр.
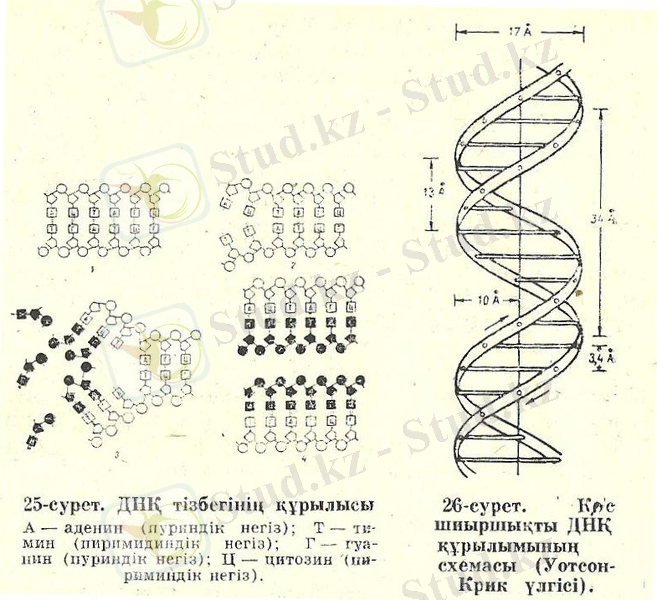
1-сурет. ДНҚ тізбегінің құрылысы
ДНҚ-ның екі тізбегін бір-бірінен ажыратуға болады. Ерітіндідегі ДНҚ молекуласын 100 С температурада қайнатса оның тізбектері бір-бірінен ажырайды, яғни олардың арасындағы сутегілік байланыстар үзіледі. Мұны денатурация деп атайды. («денатурация»-табиғи формасын жоғалту) . Құрамында ДНқ бар ерітінді қайнатып, кейін тез суыта, ажырасқан екі тізбектер жеке-жеке күйінде қалып қояды. Ал оны өте баяу суытса, ол тізбектер қайтадан бір-бірімен жұптасып, бастапқы қос тізбекті спираль түріне келеді. Жайлапсуытқан кезде тізбектерге сәйкес негіздер бірін-бірі біртіндеп тауып, қайтадан сутегілік байланыс құрай бастайды, ал тез суытқанда олар байланысып үлгере алмай қалады. Әр түрлі ДНҚ молекулаларының тізбектерінің қаншалықты оңай денатурацияланатыны құрамындағы А-Тмен Г-Ц жұптарының мөлшеріне байланысты. Жоғарыда айтып кеткендей, бір тізбектегі гуанин екінші тізбектегі цитозинмен үш сутегілік байланыс, ал аденин мен тимин екі сутегілік байланыс құрайды. Яғни, Гмен Ц-ның арасындағы байланыс А мен Т-ның арасындағы байланысқа қарағанда байланыс А мен Т-ның арасындағы байланысқа қарағанда әлдеқайда берік. Егер ДНҚ тізбектерінде Г мен Ц-ның мөлшері көп болса, онда молекуланың тізбектері тез ажырай қоймайды, ол үшін температураны неғұрлым жоғарлату керек, ал А мен Т жұбы көп болса, оның тізбектері төменгі температурада-ақ ажырай бастайды. Мысалы, өте ыстық ортада өмір сүре алатын термофильдік микроорганизмдердің ДНҚ молекуласында Г-Ц жұбының мөлшері өте жоғары болады[6] .
Әр түрлі организімдерден алынған ДНҚ-лардың бір-бірімен қаншалықты ұқсас екенін, немесе бір организмнен бөлініп алынған ДНҚ фрагментіне (мысалы, геннің) ұқсас ДНҚ бөлігі басқа организмде бар-жоғын молекулалық гибридизация әдісі арқылы зерттеуге болады. Ол жаңағы айтылған ДНҚ-ның денатурациялануына негізделген. Мысалы, өсімдіктен аса маңызды ДНҚ фрагментін (айталық генді) жануар ДНҚ-сына бар-жоғын тексеруге болады. Ол үшін өсімдікке құрамында радиоактивті атомы бар қоректік зат береді. Содан соң ол өсімдіктің ДНҚ-сынан қажетті фрагментті бөліп алады. Ал жануардан кәдімгі түрдегі (радиоакивті емес) ДНҚ-сын бөліп алып, оны шағын фрагменттерге дейін арнаулы ферменттермен үзіп, жеке тізбектерге бөледі. Осындай жануар ДНҚ-сының фрагменттерін түгелдей арнайы матрицаларға (қалыптарға) жалғастырады. Содан соң матрица түйіршіктері арқылы өсімдіктің радиоктивті фрагментін өткізеді. Егер жануардың матрицаға біріккен фрагменттерінің ішінде құрамы өсімдіктікімен толық ұқсас түрі болса онда ол өсімдік ДНҚ-сының фрагментімен қоса тізбек құрып бірігеді. Яғни, радиоактивті фрагмент матрицада қалып қояды, ал радиоактивті өсімдік фрагменті матрицада қалмаса, оған сәйкес келетін фрагменттің жануар ДНҚ-сында болмағаны. Осылай әр түрлі организмдерден алынған ұқсас ДНҚ бөлшектерінің бір-бірімен қос тізбек құруын молекулалық гибридизация деп атайды. Осы жолмен ДНҚ мен РНҚ тізбектеріндегі ұқсас бөліктерін де анықтауға болады[7] .
1. 2 ДНҚ-ның екі еселенуі (репликациясы)
Кез келген клетка бөлінер алдында оның ДНҚ молекуласы екі еселенеді және соның нәтижесінде ұрпақ клеткалары алғашқы аналық клеткадағыдай ДНҚ молекуласына ие болады. Олай болса, бөлінетін клетканың ДНҚ-сы дәл өзіне ұқсас тағы бір ДНҚ молекуласын қалай жасайды?
1940 жылы Л. Полинг пен М. Дельбрюк ген (ДНҚ) өзінше бір бейненің қалыбы секілді, ол қалыпқа саз балшық құйып, оның формасын алуға, содан кейін осы формадан қалып етіп пайдаланған алғашқы форманы қайтадан жасауға болады деген пікір айтқан. Яғни, бұл геннің алғашқы құрлымына комплементарлы ДНҚ құрылымы жасалады, одан алғашқы құрылымға сәйкес ДНҚ пайда болады деген сөз. Шынында да ДНҚ-ның бір тізбегін бір бейне десек, оған комплементарлы екінші тізбек оның кері бейнесі болып табылды. Демек, Уотсон мен Крик көрсеткен ДНҚ-ның екі тізбегі толық ажырап болғанда, екі жаңа спираль да жасалып біттеді. Алғашқы ДНҚ тізбегі ажырамай тұрғанындағы екінші ескі тізбегіне толық ұқсас болды.
 Әрине, бұл процесті де клеткадағы ферменттер жүргізеді. ДНҚ тізбектерінің бағыттары қарама-қарсы екені белгілі. Жұмысына өте мұқият ферменттер жаңа тізбекті тек бір бағытта, яғни 5-3 бағытында ғана жасайды. Олай болса, ферменттер ажыраған тізбектердіңбіреуінің бойымен жаңа тізбекті жоғарыдан төмен қарай, ал екіншісінің бойымен төменнен жоғары қарай синтездейді. Ең қызығы жаңа тізбектер үздіксіз жасалмайды, ескі тізбектің бойында бірінін кейін бірі шағын ДНҚ фрагменттері пайда болып отырады. Ондай фрагменттердің ұзындығы қарапайым бактерияларда 200 нуклеотидтен тұрса, күрделі организмдерде ол 2000-ға жуық. Осындай фрагменттерді алғаш байқаған жапон ғалымы Р. Оказаки, сондықтан оларды
оказаки
фрагменттер
і деп атайды[8] .
Әрине, бұл процесті де клеткадағы ферменттер жүргізеді. ДНҚ тізбектерінің бағыттары қарама-қарсы екені белгілі. Жұмысына өте мұқият ферменттер жаңа тізбекті тек бір бағытта, яғни 5-3 бағытында ғана жасайды. Олай болса, ферменттер ажыраған тізбектердіңбіреуінің бойымен жаңа тізбекті жоғарыдан төмен қарай, ал екіншісінің бойымен төменнен жоғары қарай синтездейді. Ең қызығы жаңа тізбектер үздіксіз жасалмайды, ескі тізбектің бойында бірінін кейін бірі шағын ДНҚ фрагменттері пайда болып отырады. Ондай фрагменттердің ұзындығы қарапайым бактерияларда 200 нуклеотидтен тұрса, күрделі организмдерде ол 2000-ға жуық. Осындай фрагменттерді алғаш байқаған жапон ғалымы Р. Оказаки, сондықтан оларды
оказаки
фрагменттер
і деп атайды[8] .
Реджи Оказаки
(1930-1975)
1953 ж. Дж. Уотсон және Ф. Крик ұсынған ДНҚ құрылымының үлгісі (моделі) генетикалық хабардын (шартты қысқарту), мутациялық өзгергіштіктің және гендердің көшірмесінің (ДНҚ молекуласының бөліктері) алынуын түсінуге мүмкіншілік берді. 1957 ж. М. Мезельсон мен Ф. Сталь, Дж. Уотсон және Ф. Криктің бактериялық клеткадағы ДНҚ-ның жартылай консервативті түрде екі еселенуі (репликация) жөніндегі көзқарасын көзқарсын дәлелдеді (27-сурет) . Ал Г. Стент ДНҚ екі еселенуінің үш түрін ұсынды: 1) консервативтік (лат. «консервативус»-сақтаушы, негізгі қалпын сақтау) еселенудеұрпақтық ДНҚ-ларда аналық ДНҚ-ның материалы болмайды; 2) жартылай консервативтік түрінде ДНҚ-ның жаңа молекуласының бір тізбегі аналық ДНҚ-дан болса, екіншісі-жаңадан құрылған тізбек; 3) дисперсиялық лат. «дисперсис»-шашырау, бытыраңқы) түрінде аналық ДНҚ-ның материал кездейсоқ шашырап жаңа ДНҚ молекуласында орын алады.
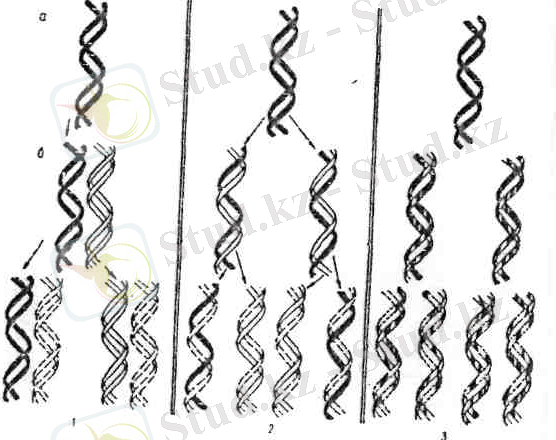
2-сурет. ДНҚ молекуласының екі еселенуі
М. Мезельсон мен Ф. Стальдың зерттеулері осы үшеуінің ішінен ДНҚ-ның жартылай консервативті екі еселену түрін таңдап алуға көмектесті. ДНҚ екі еселенуінің жартылай консервативті жолмен жүруін дәлелдеу Дж. Уотсон мен Ф. Криктің жасаған ДНК молекуласының үлгісінің дұрыстығының айғағы болды[1] .
Сонымен ДНК еселенуі оның тізбектерінің ажырауынан басталады дедік. Ол тізбектерді геликаза (хеликсспираль) -дезоксирибонуклеаза ферменттері-ДНҚ молекуласының бойымен екі бағытта жоғары және төмен ажыратады. Нуклеотидтер парлары мен ДНК-ның шиыршықты тізбегінің арасындағы сутегінің байланыстары молекуланың бір жақ шетінде бірте-бірте үзіле бастайды және (ДНҚ) тізбектердің екеуі де бірінен бірі босай отырып, жазылады. Осылайша жазылған тізбек, өзінің қосылыстарын оське тік «қоя» отырып, дезоксирибоза және фосфор қышқылының қалдықтары арасында байланыстар арқылы ұсталып тұрады. Қоршаған ортадан клеткада жинақталған бос нуклеотидтер бар, олар ДНҚ-ның жазылған тізбегінің бос қосылыстарымен реакцияға түсе алады. Бірақ әр қосылысқа бір пар, «толықтыра түсетін» қосылысы бар нуклеотид қана жуықтап, жалғаса алады. Бұл жазылған тізбекке басқа, ДНҚ-ның жетіспейтін тізбегі жалғаса бастайды деген сөз. Осы процестердің нәтижесінде ДНҚ-ның екі молекуласы пайда болды, олардың әрқайсысында қайтадан жинақталған молекуламен толықтырылған аналық молекуланың жартысы болады. Сонымен туынды молекулалар ДНҚ-ның аналық молекуласына мейілінше ұқсас келеді. Мұнда генетикалық материалдың құрамыда сақталады. Тізбектердің ажырауы мен қосылуы ферменттердің ықпалымен жүреді. Ажыраған тізбектерде оказаки ферменттері жасала бастайды. Әр фрагмент он шақты нуклеонтидтен тұратын РНҚ түріндегі жаңа тізбекті праймаза (РНҚ-полимераза) ферменті ғана бастай алады. Тізбекті бастаған РНҚ бөлшегінен ары қарай «ДНҚ-полимераза-3» деген фремент ажыраған ДНҚ бөлігіне сәйкес етіп оказаки ферментін синтездейді. Содан кейін басқа «ДНҚ-полимераза 1» ферменті фрагменттердің бастаушысы болған әлгі РНҚ тізбегін ыдыратып жібереді. Енді кезек «ДНҚ-лигаза» деген ферментке келеді. Ол оказаки фрегменттерінің арасын ескі ажыраған тізбекке сәйкес келетіндігін тексереді. Егер қандай да бір нуклеотид өз орнында тұрмаса соңғы аталған фермент оны кесіп алып тастап, оның орнына тиісті нуклеотиді қояды. Осындай әр түрлі қызмет атқаратын ферменттердің үйлесімді жұмыс жасауы тұқымдық белгінің ДНК арқылы ұрпақтарға дұрыс берілуін қамтамасыз етеді. Міне, геннің еселенуі немесе репликация дегеніміз осы[4] .
1. 3 ДНҚ молекуласының полиморфизмі
Көп уақытқа дейін ДНҚ-ның құрылымының үш түрлі қалыпы белгілі болған, олар қолайлы жағдайларда өзара өзгеру арқылы құбылып отырған. Осы үлгілердің жалпы қасиетттері төмендегі кестеде жинақталған.
1-кесте
ДНҚ құрылымдық топтар ретінде бірнеше типке кездеседі
А
В
С
Z
11
10
9 1 / 3
12
+32, 7
+36, 0
+38, 6
-30, 0
2, 56
3, 38
3, 32
3, 71
23
19
19
18
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz