Клетканың құрылымы мен қызметі: өсімдік және жануар клеткаларының құрылымдық-функционалдық ерекшеліктері

Мазмұны
Кіріспе . . . 3
І. Негізгі бөлім
- Клетка - тірі организмдің құрылымдық және функциональдық бірлігі . . . 4
- Өсімдіктер және жануарлар клеткасының айырмашылығы . . . 6
ІІ. Бөлім
Клетканың жеке бөліктерінің құрылысы мен атқаратын қызметі
2. 1 Клетка қабығы . . . 8
2. 2 Плазмалеммагеннің ыдырауы . . . 11
2. 3 Гольджи аспабы . . . 13
2. 4 Эндоплазмалық тор . . . 14
2. 5 Вакуольді жүйесі . . . 16
2. 6 Лизосомаоар . . . 18
2. 7 Клетка ядросы . . . 20
2. 8 Рибосомалар . . . 23
2. 9 Митохондриялар . . . 24
2. 10 Пластидтер жүйесі . . . 26
2. 11 Клеткалық центрі . . . 28
Қорытынды . . . 30
Қолданылған әдебиеттер тізімі . . . 31
Кіріспе
Цитология жеке ғылым ретінде өткен ғасырдың соңы ширегінде пайда болған. Бірақ клетка жөніндегі ілім XVII ғасырдан басталады, оның тарихы үш ғасырды қамтиды. Организмнің клеткалық құрылысының ашылуы микроскоптың шығуына байланысты.
Ағылшын математигі, физигі және механигі Роберт Гук 1665 жылы өзі жасаған микроскоп арқылы тығынның құрылысын қарап, оның ұяшықтардан тұратынын анықтаған. Осы ұяшықтарды клетка деп атаған. Сонымен Р. Гук : “клетка” деген терминді қалдырған. Бұл қазіргі түсініктегі клетканың ашылуы емес. Кейінірек, осы XVII ғасырда Грю мен Мальпиги микроскопты қолдана отырып өсімдіктердің құрылысын зерттеген.
XVII ғасырдың ұлы микроскопшілерінің бірі А. Левенгук, бірақ та ол өз жаңалыктарының маңызына түсіне алмаған. А. Левенгук жануарлардың клеткаларында - эритроциттерді, спермотозоидтарды, бірклеткалы жануар-ларды бірінші болып көрген. Сонымен XVII ғасырда өсімдіктердің "клеткалық құрылысы" ашылған.
Қазіргі кездегі клеткалық теорияның негізгі қағидалары мыналар:
1. Клетка тіршіліктің ең кішкене бірлігі.
2. Түрлі организмнің клеткалары құрылысы жағынан ұқсас.
3. Клеткалар бөліну арқылы көбейеді.
4. Көп клеткалы организмдер клеткалар мен олардың туындыларының жиынтығы.
XIX ғасырдың соңғы ширегінде цитология өз алдына жеке ғылым болып қалыптасты. Бұған себеп болған микроскоптың жақсартылуы мен микроскопиялық техниканың дамуы. Атап айтқанда микроскоптың штативі жақсартылып микрометрлік винт пен кремальерамен жабдықталынды. Осымен қатар микроскотың оптикасы жақсартылды.
І. Негізгі бөлім
- Клетка - тірі организмдің құрылымдық және функциональдық бірлігі.
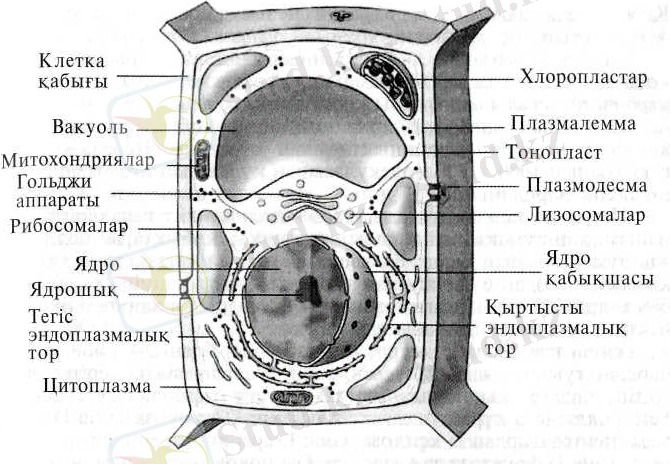
Жануарлар клеткасы плазмалық мембранамен шектелген ядро мен цитоплазмадан тұрады. Цитоплазма негізгі заттан (гиалоплазмадан), органоидтардан және кірінділерден түзіледі. Кірінділер немесе параплазмалық құрылымдар дегеніміз клеткалық метаболизмге қатыспайтын, клеткалардың көпшілігінде байқалатын алмасу өнімдерінің жиынтығы. Бұған жататындар: майдың тамшылары, гликогеннің түйіршіктері, белоктардың кристалдары және пигменттік кірінділер. Органоидтар клеткалардың барлығына дерлігінде кездеседі және заттар алмасуында маңызды функция атқарады. Бұларға эндоплазмалық тор, Гольджи аппараты, митохондриялар, тағы басқалары жатады. Клеткалардың көпшілігі бір ядролы, бірақ та екі немесе көп ядролы клеткаларда болады. Көп клеткалы организмнің клеткаларының саны өте көп. Мысалы, адам миының үлкен ми сыңарларындағы клеткалардың саны 20 млрд шамасында, ал қандағы эритроциттердің саны 25 миллиондай.
Өсімдіктер мен жануарлар организмі клеткаларының атқаратын қызметтері әр түрлі болғандықтан, олардың пішіні түрліше болады. Клеткалардың пішіні оның организмдегі орнына да байланысты. Жануарлар клеткалары жұлдыз тәрізді (нерв клеткасы), цилиндр, куб тәрізді (эпителий клеткалары), сопақша (эритроциттер), тармақталған (мезенхиманың клеткалары), жұмыр (жұмыртқа клеткасы) т. т. болады. Клеткалар пішіні жағынан ғана емес, үлкендігі жағынан да ажырайды. Мысалы, адамның жұмыртқа клеткасы (диаметрі 0, 1 мм) көлемі жағынан адамның сперматозоидынан 1 млн есе үлкен.
Адамның нерв клеткасының аксонының ұзындығы 1 м-ден артық, эпидермистің, бүйректің, бауырдың, шектің клеткаларының үлкендігінің диаметрі 30 мкм. Адамның ең кішкене клеткасы кіші лейкоциттер, диаметрі 3-4 мкм. Бактериялық клеткалар ете ұсақ келеді. Кейбірелерінің үлкендігі 0, 2-0, 3 мкм. Белгілі клеткалардың ішіндегі ұсағы микоплазманың клеткасы (0, 10-0, 25 мкм) .
Құрылысының күрделілігіне қарай клеткаларды екі топқа бөледі: прокариоттық (гректің про-алдымен, карион - ядро) деп прокариоттарға эубактериялар, көк жасыл балдырлар, спирохеталар, риккетсиялар және миколплазмалар жатады, қалған барлық организмдердің клеткалары, саңырауқұлақтардың, қарапайымдардың, жануарлар мен өсімдіктердің клеткалары эукариондық клеткалар. Прокариоттардың клеткалары ұсақ болады (0, 5-3 мкм) . Митохондриялар, хлоропластар, Гольджи аппараты; лизосомалар сияқты айқын байқалатын мембранамен шектелген органеллалар мен ядролық мембрана болмайды. Прокариоттардың гендік информацияның құрамында эукариоттар клеткасының хромосомаларында болатын негізгі белоктар - гистондар жоқ, тұйық сақина пішінді ДНК-ныд тізбегінен тұратын бір хромосомда орналасқан.
Прокариоттардың ядросында митоздық аппарат пен ядрошығыда жоқ. Прокариондық клеткалар аминқышқылдары мен көмірсулардан тұратын клеткалық қабырғамен қоршалған. Олардың плазмалық мембранасы мезосомалар деп аталатын цитоплазмаға қарап ойындылар құрайды. Прокариоттардың клеткаларына цитоплазмалық ағыс пен амеба тәрізді қозғалыс тән емес, жабайы талшықтардың көмегімен қозғалады. Прокариондық клеткалардың цитоплазмасы біртекті емес, онда фибриллалар, ұсақ мембраналық көпіршіктер мен жалпақ қапшықтар, рибосомалар, майдың тамшылары, полисахаридтер мен полифосфаттар жиынтықтарын байқауға болады. Бұлардың бәрі цитоплазманың негізгі затында, матриксінде, гиалоплазмада орналасқан.
Эукариоттардың клеткалары әдетте ірі және құрылымы жағынан прокарирттардың клеткаларына қарағанда, анағұрлым күрделі келеді. Сонымен бірге эукариоттарда жақсы жетілген ішкі мембраналар жүйесі -эндоплазмалық тор мен Гольджи аппараты, мембраналармен шектелген органеллалар - ядро, митохондриялар, хлоропласталар мен лизосомалар болады. Эукариондық клеткаларға цитоплазманың қозғалысы мен митоздық ұршық және талшықтар сияқты энергияны трансформациялаушы көптеген күрделі жүйелер тән. Эукариондық клеткалардың ядросында ядрошықтар мен хромосомалар болады.
- Өсімдіктер және жануарлар клеткасының айырмашылығы
Клеткалық қабырға әсіресе өсімдіктер клеткаларында жақсы жетілген. Саңырауқұлақтарда клеткалық қабықша хитин мен полисахаридтен тұрады, ал балдырларда, оомицеттерде және жоғарғы сатыдағы өсімдіктерде клетка-лық қабырғаның негізгі компоненті полисахарид, целлюлоза. Клеткалық қабықшаның гликопротеидтері мен полисахаридтері клетканың ішіндегі Гольджи аппаратының қатысында синтезделеді де, бірнеше қабаттан тұратын күрделі клеткалық құрылым күйінде клеткадан экзоцитоз арқылы сыртқа шығарылады.
Жануралар мен өсімдіктер клеткаларының үш негізгі құрылыстық айырмашылықтары бар: 1) жануарлар клеткаларында болатын центриоль жоғары сатыдағы өсімдіктер клеткаларында болмайды; 2) өсімдіктер клеткаларының жануарлар клеткаларынан өзгешелігі цитоплазмасында пластидтер болады; 3) өсімдіктер клеткаларына целлюлозадан тұратын қатты клеткалық қабырға тән.
Сыртқы ортадан алынатын энергияның түріне байланысты тірі клеткаларды үлкен екі типке бөлуге болады. Бірінші типтің клеткаларын гетеротрофты клеткалар деп атайды. Бұған жататындар: адам организмінің барлық клеткалары мен жоғары сатыдағы жануарлардың клеткалары. Бұл клеткаларға химиялық құрамы күрделі дайын органикалық молекулалардың үнемі келіп тұруы қажет. Атап айтқанда, көмірсулар, белоктар мен майлар.
Гетеротрофты клеткалар энергияны осы күрделі заттарды тотықтыру арқылы алады. Гетеротрофты клеткалар бөлінген осы энергияны өзінің биологиялық функцияларының қажетіне жұмсайды.
Автотрофты немесе фотосинтездеуші клеткалар клеткалардың екінші типін құрайды. Бұған жататындар: жасыл өсімдіктердің клеткалары. Фотосинтез процесі кезінде олар күн сәулесінің энергиясын хлорофилл пигментінің көмегімен химиялық энергияға айналдырады және атмосфералық көміртегінің қос тотығынан көміртегін бөліл алып, жабайы органикалық молекуланы (глюкозаның молекуласын) түзеді. Жасыл өсімдіктер мен басқа организмдердің клеткалары өзінің құрамына кіретін күрделі молекулаларды да құрайды. Сонымен барлық тірі организмдер энергияны күн сәулесінен алады; өсімдік клеткалары энергияны тікелей күннен алатын болса, ал жануарлар оны жанама жолмен алады.
ІІ. Бөлім
Клетканың жеке бөліктерінің құрылысы мен атқаратын қызметі
2. 1. Клетка қабығы
Өсімдік клеткаларының пішінін, көлемін анықтайтын және оларды бір-бірінен ажырататын, сыртын қоршаған, негізінен полисахаридтерден құралған, тығыз қабығы болады. Ол клетканың митоздық бөлінуінің телофазалық кезеңінде қалыптасады.
Ұрықтық ұлпалардың және клеткалардың созылып өсу кезеңіндегі қабығын алғашқы (бастапқы) деп атайды. Кейінірек осы қабықтың ішкі (астар) жағынан қайталама қабық қалыптасады. Осыған байланысты қабық қалындап, қатайып беріктенеді.
Алғашқы қабықтың құрамына енетін бөліктерін төрт топқа бөлуге болады: 1. Құрылыстық бөліктер - целлюлоза (автотрофты өсімдіктер), хитин (саңырауқұлақтарда), глюкан (ашытқыларда), маннан немесе ксилан (кейбір балдырларда) . 2. Қабық негізінің (матрикс) бөліктері - гемицеллюлозалар, пектиндер, белоктар, липидтер. 3. Қабықтың астарын лигнин, суберин және басқа қосындылар бедерлейді. 4. Қабықтың тыс жағының құрамына кутин және балауыз енеді.
Клетка қабығында, әсіресе эпидерма клеткаларында, аталған қосындылардан басқа, біршама мөлшерде кальций силикаты мен карбонаты сияқты минералды қосындылар да кездеседі.
Клетканың алғашқы және қайталама қабықтарының негізгі қаңқасы целлюлоза талшықтарынан құралады. Целлюлоза - Д-глюкозаның 1000-11000 молекуласынан құралатын өте ірі молекулалы полисахарид.
Шамамен, целлюлозаның 100-200 полимерлі тізбегі бірігіп қарапайым жіпшумаққа айналады. Алғашқы қабықтағы целлюлоза жіпшумақтарының көлденең өлшемі 4 нм-дей ғана болса, қайталама қабықта ол одан 6 есе жуандау болады. Осы жіпшумақтар бірімен-бірі матрикс құрамындағы гемицеллюлозалар, гликопротеидтер және пектиндер арқылы байланысады.
Клетка қабығының құрылысы. Жалпы құрамының күрделілігіне қарамастан қабықтың қалындығы 0, 1 ден ондаған микрон аралығында болады. Меристеманың бөлінетін және созылып өсетін клеткаларын қоршаған алғашқы қабық біртіндеп қалыңдап, қайталама қабыққа айналады. Көрші екі клетканың алғашқы қабықтарының арасында табақша (пластинка) деп аталатын аморфты үшінші қабықша пайда болады. Жалпы клетка қабығының құрамына енетін бөліктердің өзара байланысу жолдары 1. 2-суретте бейнеленген. Онда, ең алдымен целлюлоза молекулалары өзара сутекті байланыспен жалғасып жідішке жіпшумақтарға бірігеді. Оларға гемицеллюлозалар да сутек арқылы байланысады. Клетка қабығындағы пектин, гемицеллюлоза және экстенсин (белок) молекулалары өзара коваленттік байланыс арқылы тұтасады. Пектинді полисахарид - ксисоглюканның бір тізбегі целлюлозаның бірнеше тізбегімен байланысса, целлюлозаның бір тізбегі ксилоглюкан арқылы бірнеше пектинді полимерлермен байланысады. Сөйтіп қабықтың құрылысында өзара айқасқан коваленттік байланыстар жүйесі пайда болады.
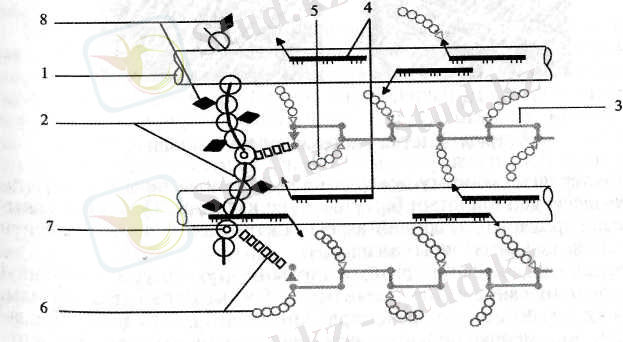
1. 2-сурет. Алғашқы қабық бөліктерінің өзара байланысу желісі
(Bauer, 1977) :
1 - целлюлоза жіпшумағы; 2 - белок; 3 - рамногалактуронан; 4 -силоглюкан; 5 - 4-байланысқан (арабино) галактан; 6 - 3-байланысқан (арабино) галактан; 7 - экстенсиндегі серин қалдықтары; 8 - экстенсин тетрарабинозиді.
Бұл суреттегі ұлгі қос жарнақты өсімдіктер клеткасының алғашқы қабық құрылысын көбірек бейнелейді. Бірақ оны дара жарнақты өсімдіктерге де тән деуге болады. Себебі, олардың клетка қабығындағы негізгі бөліктердің ара қатынастары қос жарнақтыларға ұқсас екендігі анықталды.
Меристемалық клеткалардың алғашқы қабығында жіңішке жіпшумақтар қабық негізінде (матрикс) пішінсіз тор жасап орналасады. Клетка ұзарған сайын жіпшумақтардың орналасуы кәдімгі тузу пішінді тордың құрылысын бейнелей бастайды. Алғашқы қабықтың астар жағындағы жіпшумақтар ұзарып, өсу бойына көлденең бағытталып, тыс жағындағылар қатарласа орналасады. Қабық бөліктерінің бұндай орналасуын көп қабатты торлы құрылыс деп атайды.
Көптеген клеткалардың алғашқы қабығында соңдай құрылысты аймактардың аралықтарында ұзарып өсу бағытын бойлай жайғасқан жіпшумақ тартпалар пайда болады.
Әртүрлі клеткалардың алғашқы қабығындағы бөліктердің ара қатынастары түрліше болып келеді. Мысалы, талшық клеткаларында көлденең тартпалар болмайды да, қабық біртұтас көп қабатты торға ұқсаңды; паренхималық клеткаларда осы бөліктер бірдей кездеседі; колленхималық клеткаларда көлденең тартпалар басымырақ болады.
Ұлпалардағы және мүшелердегі клетка қабықтары өзара ұштасып апопласт деп аталатын біртұтас жүйе құрайды. Осы жүйе арқылы клетка араларында, сондай-ақ өсімдіктің жалпы денесінде еріген заттар оңай, кедергісіз тасымадданады.
Клетка қабығындағы ерігін заттардың ірігуі, сіңуі және бөлінуі жүзеге асатын саңылаулар жиынтығы "бос кеңістік" деп аталады. Клетка қабығыңда полисахаридтерді синтездейтін ферменттер болмағанымен, полимерлердің өзгеру реакцияларына қатысатын ферменттер бар екеңдігі анықталды. Оларға α - және β-глюкозидазалар, α-және β- галактозидазалар, маннозидазалар, целлюлаза, β-фруктофуранозидаза, полигалактоуроназа (пектиназа), пектинэстараза, глико-зилтрансфераза жөне т. б. жатады. Сонымен қатар клетка қабығында пероксидаза, аскорбатоксидаза және фосфатаза ферменттерінің де болатындығы байқалды.
Қос жарнақты өсімдіктердің клетка қабықтарында көптеген зиянды микроорганизмдерден бөлінетін полигалактоураназа ферментін басытқылайтын, соған сәйкес клеткаларды індеттену әсерінен қорғайтын белоктардың болатындығы да анықталды.
Өсімдіктің әр түрлерінде клетка қабығындағы полисахаридтер құрамы біркелкі болмайтындығы, ал белгілі түрге жататын өкілдерде олардың өзара ұқсас екендігі дәлелденді.
Клетка қабығының қалыптасуы полимерлердің пайда болуынан басталып, клетка және өсімдік өмірінің кезендерінде түрлі өзгерістерге ұшырап, күрделі құрылымға айналуымен аяқталады. Бұл процеске клетка ядросы, рибосомалар, эндоплазмалық тор, Гольджи аппараты, плазмалеммалар қатысады.
2. 2. Плазмалемма
Клетка қабығының астар жағындағы жұқа қабықша плазмалемма деп аталады. Оның қалыңдығы 7, 5-10, 0 нм шамасында болады. Плазмалемма қабықсыз клеткаларды сыртқы ортамен, қабықты клеткаларды өзара және цитоплазмамен байланыстырады. Цитоплазмадағы басқа мембраналар сияқты белокты және қос липидті екі қабаттан тұрады.
Оның тыс жағында қабықты қалыптастыруға қатысатын ферменттер болады деген болжамдар бар.
Плазмалемманың құрамыңда липидтердің мөлшері - 40-54%. Оның ішінде стериндер мен бейтарап липидтер 30-50%. Басқа мембраналармен салыстырғанда, плазмалеммада қаныққан май қышқылдары көбірек және сыртқы қабатында гликолипидтер басымдау орын алады.
Плазмалемманың су өткізгіштігі жоғары. Липидті қабатында пиаметрі 0, 4 нм шамасыңда, ерекше су өткізгіш молекулалық саңылаулары болады деген мәліметтер кездеседі. Белокты бөлігі липидтермен қосылып плазмалемманың негізгі қызметтерін қамтамасыз етеді. Мембрана арқылы заттардың тасымалдануы белокты жүйелер арқылы жүзеге асады. Олардың өкілдері ретінде, балдырлардағы, цитоплазмадан Н + және НСО 3 - иондарын ортадағы К + және С1 - -иондарына алмастыратын бейтарап иондық насостарды және өсімдік плазмалеммаларында болатын протондық насостарды атауға болады. Протондық насостың қызметі АТФ молекуласын гидролиздейтін Н + АТФазаға негізделген. Клеткадағы басқа қосындылардың, мысалы, қанттар мегі амин қышқылдардың тасымалдануы да плазмалем-мадағы Н + насостардың ырықтығына байланысты.
Плазмалемманың синтездік функциясы полисахаридтердің синтезделуіне қатысатын белоктық жүйелерге, яғни 1, 3-р-глюкансинтаза және целлюлозосинтазаларға байланысты.
Сонымен қатар плазмалеммада ауксин және цитокинин сияқты қоздырғыштар (рецепторлар) да болуы мүмкін.
Бактериялардың плазмалеммаларында тыныс алу тізбегі орналасқан. Жоғары сатыдағы өсімдіктер плазмалеммаларында электрондардың, флавопротеинді оксиредуктазаның және цитохромның (b 5 ) қатысуымен жүзеге асатын тасымалдау тізбегі болатындығы анықталды.
Плазмалемма өсімдік организміндегі зәр бөліну (секреция) процестеріне де қатысады. Плазмалемманың жартылай өткізгіштігі, құрылысы метаболипік процестермен реттеледі.
Сонымен плазмалемма клеткада заттардың енуі мен бөлінуін реттейді, клетка қабығының қалыптасуына қатысады, рецепторлық және осмостық процеске байланысты қызметтер атқарады.
Клеткадағы плазмалемма үздіксіз қозғалыста болады және қайталанып қалыптасып жатады.
2. 3. Гольджи аспабы (аппараты, ГА)
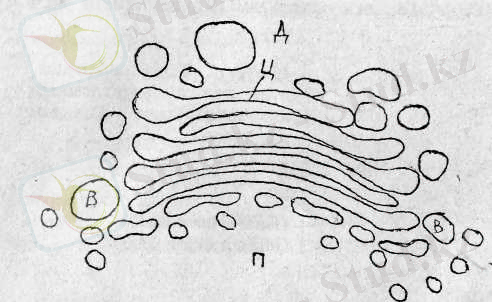
Камило Гольджи (1899) ашқан клетканың бұл органоиды диктиосома, везикула (көпіршік, бүртік) және цистерна (диктиосома) аралық түтікшелер сияқты құрылымдардың жиынтығы екендігі анықталды.
Қысыңқы цистерналар - диктиосомалар (гольджисомалар) 5-8 да-надан бірігіп буда сияқты топтасып орналасады. Олардың қоршау мембраналарының қалыңдығы 7-8 нм шамасында болады.
 Диктиосома
мембраналарыңда көптеген ферменттік жүйелер ашылды: 1. Клетка қабығының матрикстік (негіздік) құрамына енетін полисахаридтерді (пектин, гемицеллюлоза) синтездеуге қатысатын ферменттер; 2. Клетка қабығындағы экстенсинді (белок) гликозилдейтін - УДФ арабиноза: арабинозилтрансфераза; 3. Балдырлар клеткасыңдағы полисахаридтерді сульфаттауға қатысатын ферменттер; 4. Гликозилпротеидтер мен гликолипидтер пайда болған кезде белоктарды және липидтерді гликозилдейтін УДФ галактоза: N-ацетилглюкоза- мен УДФ глюкоза: ; 5. Диктиосома мембраналарын өзгертуге (модификациялауға) қатысты трансглюкозидаза, т. б.
Диктиосома
мембраналарыңда көптеген ферменттік жүйелер ашылды: 1. Клетка қабығының матрикстік (негіздік) құрамына енетін полисахаридтерді (пектин, гемицеллюлоза) синтездеуге қатысатын ферменттер; 2. Клетка қабығындағы экстенсинді (белок) гликозилдейтін - УДФ арабиноза: арабинозилтрансфераза; 3. Балдырлар клеткасыңдағы полисахаридтерді сульфаттауға қатысатын ферменттер; 4. Гликозилпротеидтер мен гликолипидтер пайда болған кезде белоктарды және липидтерді гликозилдейтін УДФ галактоза: N-ацетилглюкоза- мен УДФ глюкоза: ; 5. Диктиосома мембраналарын өзгертуге (модификациялауға) қатысты трансглюкозидаза, т. б.
Диктиосома құрылысының схемасы. П - проксималь бөлігі, Д - дистальдік бөлігі, В -вакуольдар, Ц -жалпақ мембраналық истерналар
Синтезделген заттар (полисахарңдтер, белоктар, липидтер және гликопротеидтер) мембранаға еніп, секретті көпіршіктер күйінде плаз-малеммаға тасымалданады. Плазмалеммамен байланысқан көпіршіктер экзоцитоздық жолмен секреттік заттарды плазма айналасындағы кеңістікке шығарады. Көпіршік мебраналары плазмалыққа айналады. Жаңа мембраналық бөліктердің қосылуынан плазмалемма жаңарып, өсуі жеделдейді. ГА мембраналық жүйесі де үздіксіз жаңарып тұрады.
Секретгік заттар бөлінген сайын ГА мембраналары плазмалемманы қалыптастырады. Олардың мембраналары онтогенездік және функциялық жағынан өзара ұқсас.
Сонымен, клеткадағы ГА мембраналарды құрастырып, полисахидтерді синтездеп, сульфаттау, белоктар мен липидтерді гликолиздеу қызметгерін атқарады. Өсімдік клеткасы онтогенезінің барлық сатыларында Гольджи аспабы қабықтың қалыптасуына қатысады. Бұл процеске аралық қабаттардың пайда болуы, клетка қабығының өсуі, матрикстік заттардың синтезделуі, қайталама қабықтың қалыптасуы, шырыштардың қабықтың және клетканың сыртына бөлінуін, т. б. жатқызуға болады.
Гольджи аспабы эндоплазмалык тор мен плазмалемма мембраналарын өзара байланыстыратын түйін болып есептеледі.
2. 4. Эндоплазмалық тор
Сыртқы плазмалық мембранамен генетикалық байланыста болатын, клетканың ішкі мембраналар жүйесін 1945 жылы Г. Палад ашып, К. Портер эндоплазмалық ретикулум (ЭР), немесе эндоплазмалық mop (ЭТ) деп атады. Ол өзекше (канал), цистерна, көпіршік сияқты құрылымдардан құралған жүйе. Олардың саны және орналасуы клетканың қызметі мен физиологиялық күйіне байланысты өзгеріп тұрады. Ядро қабықшасы да осы жүйеге қосылады. Құрылымдық пішініне қарап ЭТ кедір-бұдырлы және бірыңғай тегіс болып екі түрге бөлінеді. Біріншісінің өзекшелер жүйесінің бетінде рибосомалар орналасады да, ал тегіс түрінде олар болмайды. Осы екі түр бір-біріне ауысуы мүмкін.
Эндоплазмалық тор мембраналарындағы ферменттер эстеразалар және тотығу жүйесінің ферментгері болып екі топқа бөлінеді. Бірінші топқа глюкозо-6-фосфатаза, Mg - тәуелді нуклеозидфосфатазалар және ерекшеленбеген эстараза жатады. Кедір-бұдырлы ЭТда болатын глюкозо-6-фосфатаза мембрананың гидрофобты бөлігі арқылы глюкозаны ауыстыруға қатысуы мүмкін. Ол анорганикалық пирофосфат, маннозо-6-фосфат, нуклеозид ди- және трифосфаттардан пирофосфат тобын глюкозаға ауыстырады. Mg - тәуелді нуклеозидфос-фатазалар нуклеозидтердің моно-, ди- және трифосфорлы эфирлерін гидролиздейді. ЭТ мембранасындағы эстераза рН 6, 5-7, 5 арасында қысқа май қышқылдары мен n-нитрофенолға әсер етеді.
 Эндоплазмалық тордағы тотығу жүйесінің орналасуы митохон-дриялардағыдай - тізбек басында флавопротеидтер, ең соңында -цитохроммен бітеді. Бірақ микросомолар мембранасындағы ФАД-ты флавопротеид - НАДФН-цитохром-с-редуктаза митохондрия мембра-наларында болмайды. Екінші НАДН-ты - НАДН-цитохром-с-редуктаза ферменті тек
b
цитохромды тотықсыздандырады.
Эндоплазмалық тордағы тотығу жүйесінің орналасуы митохон-дриялардағыдай - тізбек басында флавопротеидтер, ең соңында -цитохроммен бітеді. Бірақ микросомолар мембранасындағы ФАД-ты флавопротеид - НАДФН-цитохром-с-редуктаза митохондрия мембра-наларында болмайды. Екінші НАДН-ты - НАДН-цитохром-с-редуктаза ферменті тек
b
цитохромды тотықсыздандырады.
Гранулалық эндойлазмалық тор ( х 9)
Цитохром b электронды бір мембраналық жүйеден екіншіге, мысалы, ЭТ-дан митохондрияларға ауыстыра алатындығы анықталды.
Эндоплазмалық тор клеткада алуан-түрлі қызмет атқарады. Мысалы, кедір-бұдыр ЭТ белоктарды синтездеп қорға айналуын және тасымалдануын жүзеге асырады. Сонымен қатар, белоктардың секреттенуіне (тысқа бөлінуі) де қатысады. ЭТ-дың тегіс түрі көмірсулардың, липидтердің, т. б. заттардың синтезделуіне және клеткаға уытты әсер ететін гидрофобты қосындылардың залалсыздану процестеріне қатысады. Эвдоплазмалық тор, митоховдриялармен қатар, өсімдік клеткасындағы тотығу-тотықсыздану жүйелерінің негізгі бөлігі болып есептеледі. Сондай-ақ, ол клеткадағы тітіркену әсерін таратушы және зат алмасу процестерін өзара байланыстырушылық та қызмет атқарады. Эндоплазмалық тор ядролық және плазмалық мембраналардан пайда болып, рибосомалардың қатысуымен синтезделуі, немесе митоз кезінде өзгеріске ұшырамай, аналық клеткадан ауысуы да мүмкін.
2. 5. Вакуольдер жүйесі
Өсімдік клеткаларының негізгі ерекшеліктерін бейнелейтін белгісіне вакуольдер жүйесі жатады. Сыртқы пішіні жағынан бұл қүрылым да әртүрлі болып келеді. Меристема клеткаларында өте майда көпіршіктер күйінде болатын болса, өсіп жетілген клеткаларда, олардың өзара тұтасуы нәтижесінде үлкен орталық вакуольге айналады. Көптеген жағдайда оның көлемі жалпы клетка көлемінің 90%-не жақындайды. Вакуольдік жүйенің қалыптасу механизмдері цитологиялық, электронды-микроскоптық және биохимиялық әдістерді пайдалану арқылы зерттеліп келеді. Осы әдістерді бірдей пайдалану нәтижесінде Швейцария цитологы Ф. Матиль (1968) өсімдік клеткасында вакуольдер жүйесінің қалыптасуының жолдары төмеңдегідей бағытта болуы мүмкін деген қорытындыға келді.
Вакуольдік жүйенің майда көпіршіктер сияқты алғашқы бастамалары (провакуольдер) меристема клеткаларында байқалады. Қазіргі кезде электроңдық микроскоп көмегімен провакуольдер эндоплазмалық тор цистернасының кеңуінен де пайда болатындығы дәлелденді. Дәнді дақылдардағы алейрон қабатының клеткаларында вакуольдер осылайша қалыптасады.
Клеткалар өскен сайын провакуольдер бірігіп, ірі вакуольге айналады. Оның айналасы эндоплазмалық тор мембранасының туындысы вакуольдік мембрана - тонопластпен қоршалады.
Қалыптасқан тонопластың ішке қарай доғалданып созылуының нәтижесінде цитоплазмадан вакуольге енген денешік гидролиздік ферменттердің әсерінен ыдырап кетеді.
Гольджи аппаратының көпіршік тәрізді туындылары вакуоль мембранасымен тікелей араласпастан, тонопластпен құндақталып, вакуэльге енгесін ыдырайды.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz