Фотосинтез: жарық сатысы, циклді және циклсіз электрон тасымалы, фотосинтетикалық фосфорлану және Кальвин циклі

Мазмұны
Кіріспе
1. Фотосинтездің жарық сатысы
- Фотосинтездік жүйелердегі энергия ауысу процестері
- Электрондардың циклсіз (айналымсыз) тасымалдануы және НАДФН пен АТФ-тың синтезделуі
- Электрондардың циклді тасымалдануы
- Фотосинтездік фосфорлану
- Электрон жене протовдар тасымалдану реакцияларының тилакоид мембраналарында орналасуы
2. Фотосинтездің қараңғы сатысы
2. 1 Фотосинтездің С 3 - жолы (Кальвин циіслі)
2. 2 Фотосинтездің С 4 - жолы (С 4 -дикарбон қышқылдар - Хэтч - Слэк циклі)
Қорытынды
Пайдаланған әдебиет
1. Фотосинтездің жарық сатысы
1. 1 Фотосинтездік жүйелердегі энергия ауысу процестері
Фотосинтездің жарық сатысы фотосинтездік жүйелер құрамындағы пигменттердің жарық квантын (фотон) сіңіруінен басталады. Жоғарыда көрсетілгендей жеке пигменттердің сіңіретін сәулелері спектрдің әр түрлі аймақтарына тура келеді. Бұл ерекшеліктер олардың молекулаларындағы электрондардың орналасу реттеріне байланысты.
Пигменттің фотонды сіңіруінен оның молекуласындағы элект-рондардың орналасуы өзгеріп "ырықтанған" күйге ауысады. Мысалы, қызыл сәуле квантын сіңірген хлорофилл молекуласы қозған синтглеттік - S 1 * күйге көшеді (1-сурет) . Энергиясы молырақ көк сөуле кванты сіңгенде электрон одан да жоғары қозған деңгейге (орбиталь) - S 1 * көтеріледі. Хлорофилдің қозған - энергетикалық деңгейі жоғарылаған молекуласы бірнеше жолмен бұрынғы қалпына қайтып келеді. Осы күйдегі энергияның біраз бөлігін жылу түрінде бөліп шығарып хлорофилл молекуласы ұзын толқынды жарық квантын сәулелендіруі мүмкін. Бұл құбылыс флуоресценсия деп аталады. Энергияның ендігі белігі молекула синглеттік қозған күйден тұрақтылау триплеттік - Т күйге ауысқаңда бөлінеді. Осы күйдегі молекула одан да ұзын толқынды жарық квантын сәулелендіріп (фосфоресценсия) негізгі күйіне қайтып келеді.
Ең соңында қозған молекуладағы энергия фотохимиялық реакцияларға жұмсалады.
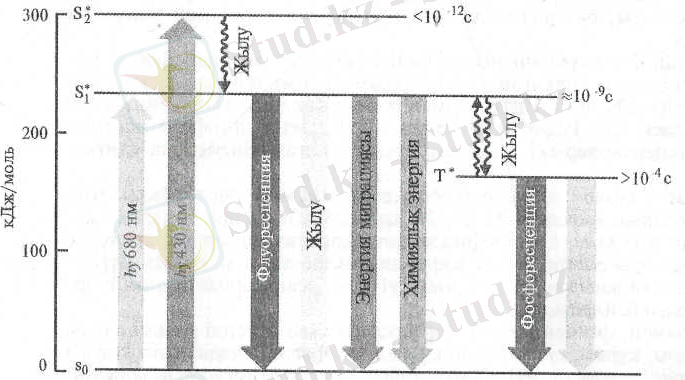
1-сурет. Хлорофилл молекуласының эиергетикалық күйлері және электроңдық қозу энергиясының жұмсалу жолдары. Сандар молекуланың жеке күйдегі өмрін көрсетеді (секунд) (В. В. Полевой, 1989) .
Осындай өзгерістер фотосинтездік жүйелер құрамындағы барлық пигмент молекулаларына тән деуге болады. Яғни, олардың әрқайсысы өздерінің молекулалық құрылысына байланысты толқын ұзындықтары түрліше сәулелерді сіңіріп, оны соңында реакциялық орталықтағы фотохимиялық реакцияға тікелей қатысатын хлорофилл (Р 700 , Р 680 ) молекулаларына жеткізеді.
Бірінші және екінші фотосинтездік жүйелердегі энергияның ауысуы жоғарыдан төмен қарай аққан сияқты болады. Оның мәнісі квант энергиясы антенналық пигменттерден реакциялық орталықтағы Р 700 және P 680 пигменттеріне жеткенше біртіндеп азаяды. Сонымен, жарық жинаушы антенна пигменттері (ЖЖП) реакциялық орталық пигменттеріне қарағанда энергиясы молырақ (қысқа толқынды) фотондарды сіңіреді. Сондықтан, қозу энергиясы фотожүйелерде Р 700 немесе Р 680 пигменттерге жеткенсін кері бағытта ауыспайды да, фотохимиялық реакциялар энергияның жұмсалуын ең тиімді жолы болып қалады.
Осы айтылғандарға сәйкес фотожүйелердегі энергия ауысуының мүмкін бірізділігін төмендегіше бейнелеуге болады (2-сурет) .
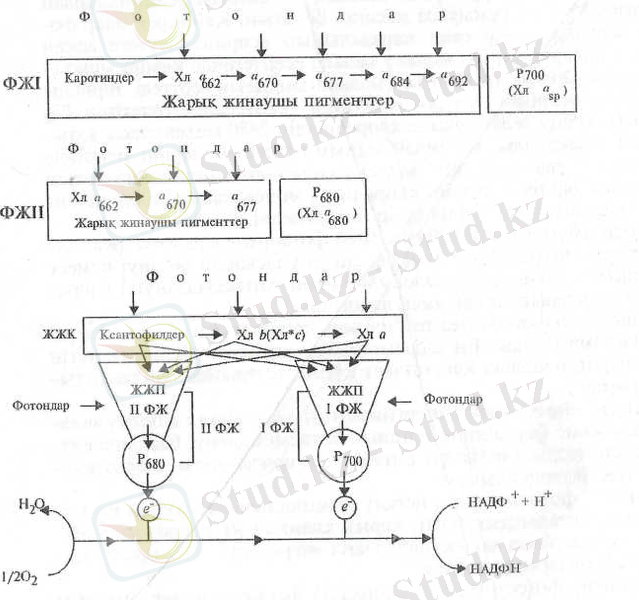
2-cypeт. Фотожүйелердегі және тиісті жарық жинаушы комплекстегі пигменттердің болжамды орналасуы және әрекеттестігі (Т. Гудвин, Э. Мерсер, 1986) :
І, II ФЖ - фотожүйелер, ЖЖК - жарық жинаушы комплекс.
Қозған пигмент молекулаларындағы энергияның I және II фотожүйелердегі Р 700 немесе Р б80 пигменттерге жеткенге дейінгі гео-метриялық жылжу жолы бағытсыз, кездейсоқ шарлауға ұқсас деп есептеледі. Реакциялық орталықтардың жарық жинаушы молекулалардың арасында орналасуы кездейсоқ болуы мүмкін. Демек, біріншіден, антеннадағы кез келген молекулаға сіңген энергия басқа молекулаларға ауысып, соңында кез келген реакциялық орталыққа енуі мүмкін. Екіншіден, белгілі бір реакциялық орталық антеннадағы молекулалардың белгілі тобынан ғана энергия қабылдап және пигментгік жүйедегі сіңген кез келген квант және тек бір ғана реакциялық орталыққа енуі мүмкін.
Реакциялық орталықтағы фотохимиялық реакция өте жылдам жүзеге асатын болуы керек. Ондай болмаса, қозған синглеттік күйдегі Р 700 және P 680 өз энергиясын флyopeсцeнцияғa шығындар еді. Шындығында фотосинтездеуші бактерияларда фотохимиялық реакция шамамен 6·10 12 секундте, яғни флуоресценция мезгілінен бірнеше рет жылдамырақ іске асады екен. Ендеше, реакция осындай жылдамдықпен өтуі үшін пигменттердің әрқайсысы Р 700 (немесе Р 680 ) тилакоид мембранасында өз энергиясын беретін тотығу-тотықсыздану жүйесімен қатар орналасуы қажет. Сондай-ақ, тотыққан Р + 700 (немесе Р + 680 ) пигментке электрон беретін тотығу- тотықсыздану жүйесі онымен қатарласып, жақын орналасуы керек.
Қозған Р + 700 және Р + 68О пигменттерінен бөлінген электрондардың әсерінен феофитин және А деп белгіленетін белокты жүйелердің тотықсыздануымен жарық қажет ететін реакциялары аяқталады. Басқа реакциялардың барлығы, соның ішінде НАДН жөне АТФ-тың пайда болуы да жарықты қажет етпейді.
Дегенмен, фотосинтездің жарықты қажет ететін (жарық сатысы) реакциялары өте күшті тотықсыздырғыш НАДФН-тың және макроэргті АТФ-тың синтезделуімен аяқталады деуге болады. НАДФ-тың НАДФН-қа айналуына (тотықсыздануына) қажетті электрондар су молекуласынан бөлінеді.
1. 2. Электрондардың циклсіз (айналымсыз) тасымалдануы және НАДФН пен АТФ-тың синтезделуі
Бұл процесс циклсіз фотосинтездік фосфорлану деп аталады, циклсіз деп аталуы - электрондардың айналымсыз жолмен тасымалдануына байланысты да, фото деп аталуы электрондар тек энергияның әсерінен жылжитындығында; фосфорлану деп аталу себебі бұл процесте бөлінген энергия АДФ-тың фосфорланып АТФ синтезделуіне жұмсалатындығына байланысты.
Электрондардың тасымалдану жолын шартты түрде 5 бөлікке бөлуге болады. Олардың екеуі пигменттік (фотосинтездік) жүйелер, үшеуі - электрон тасымалдаушы тізбектер: біреуі электрондарды қозған Р 700 -ден алып НАДФ + -қа жеткізеді; екіншісі (аралық) - электрондарды қозған күйдегі Р* 680 пигменттен Р* 700 -ге жеткізеді; оттегінің бөліну жүйесі деп аталатын үшіншісі электрондарды су молекуласынан қабылдап, Р + 680 пигментіне ауыстырады.
1. Электрондардың Р* 700 -ден НАДФ + -қа дейін тасымалдану тізбегі.
Осы тізбектегі қозған күйдегі Р* 700 пигменттен электрондарды бірінші болып қабылдайтын қабылдағыштың (акцептордың) тегі көпке дейін анықталмай, шамамен X деп белгіленіп келді. Кейінірек ол акцептордың а хлорофилдің мономерлі түрі (А, ) екендігі анықталды. Одан электрондар құрамында темір, күкірт атомдары бар белоктарға (FeS - белоктар) тасымалданады. Олардың алдыңғысы A 2 деп, соңғысы А в деп белгіленеді. Тотықсызданған FeS-белоктан (А в ) электрондар ферредоксинді жүйеге ауысады. Хлоропластардағы ферредоксин үш түрлі күйде кездесетіндігі дәлелденді: еріген ферредоксин - потенциалы Е΄ 0 =-0, 43 В - 0, 43 В болатын, құрамында (2Fe-2S) -орталығы бар; қалған екеуі - ФД 590 және ФД 540 (потенциалдары E'=-0, 59 және -0, 54) мембраналарда байланысқан, орталықтары - 4Ғe-4S. Тотықсызданған А в осылардың біреуін (яғни ФД 540 ) тотықсыздандырады да, одан электрон еріген күйдегі ферредоксинге ауысады. Ал, ферредоксиннің ФД 594 түрі электрондардың циклі (айналым-ды) тасымалдануына қатысады.
Еріген ферредоксин хлоропластардағы тотығу-тотықсыздану жүйесі - фермент: НАДФ-оксиредуктаза және ФАД-протеинмен 1:1 қатынастағы жиынтық құрамына енеді. Бұл жиынтық тилакоидтың сыртқы мембранасында әлсіз байланысқан деп есептеледі. ФАД-ферредоксин: НАДФ-редуктазаның тотықсыздануына екі электрон және екі протон қажет. Протондар қоршаған ортадағы (ұлпа) судан, электрондар - тотықсызданған ферредоксиннің екі молекуласынан алынады. Содан соң тотықсызданған флавопротеин НАДФ + -ты тотықсыздандырады. Нетижесінде НАДФН және Н + пайда болады.
2. Электрондардың аралық тасымалдану тізбегі. Электроның басқа акцепторға берген Р + 700 пигменті бос катион - радикалға (Р + 700 ) айналады. Ол бұрынғы қалпына келіп, циклсіз тасымалдану жалғасуы үшін қажетті электрондарды (энергия) потенциалы жоғарылау (Е' 0 =+0, 37В) пластоцианиннен алады. Пластоцианин -молекулалық массасы 21000 шамадағы белок. Оның хлоропластағы концентрациясы хлорофилдердің жалпы мөлшерінің 0, 2%-дей шамада болады.
Электронын Р + пигментке беріп, тотыққан пластоцианинді f цитохром (Е' 0 =+0, 36В) тотықсыздандырады. Тотыққан f цитохром, екі электронды, потенциалы Е' 0 =+0, 1В, тотығу-тотықсыздану жүйе - пластохинон f пластохинол (суретте ПХ-ПХН, ) арқылы тотықсызданады. Бұл жүйеден екі электронмен қатар екі протон да бөлінеді. Олар АТФ синтезіне қажет.
Хлоропластардағы пластохинон мөлшері басқа тотығу-тотықсыздану жүйелерінің мөлшерінен анағұрлым көбірек, концентрациясы хлорофилдің жалпы концентрациясының 5-10 %-не жетеді. Сонымен қатар осы тізбектегі тасымалдаушылардың ішінде пластохинон ғана электрон акцепторларының көптеген қорын түзеді.
Пластохинонды тотықсыздандыратын b 559 цитохром потенциалы (Е' о ) жағынан екі түрлі болады. Соның потенциалы төмендеуі арқылы пластохинон тотықсызданады. Кейбір жағдайда Р пигментті тотықсыздандыра алатын, потенциалы жоғарылау түрі (Е' о = + 0, 37) циклсіз тізбекке қатыспайды.
Төмен потенциалды феррицитохром b өз кезегінде электронды тотықсызданған Q - -ден (Q/Q") Е' 0 =-0, 04В) алады, яғни Р* 680 элект-рондарының бірінші акцепторы болып табылады.
Сонымен, аралық тасымалдану тізбегінде электрондар потенциалы Е' о = + 0, 37 В деңгейге дейін төмендейді; бұнда электрондардың екі моліне есептегенде 79 кДж энергия бөлінеді. Бұл энергия, протондардың концентрация айырмашылығы (градиент) күйінде сақталып, АДФ пен анорганикалық фосфаттан 2 моль АТФ синтезделуге пайдаланылады. Бұл айтылғаннан аралық электрон тасымалдау тізбегінің негізгі нәтижесі АТФ синтезі екендігі байқалады. Сондықтан II фотожүйенің қызметі элект-рондарды су потенциалынан (Е' о = + 0, 82 В) теріс потенциал деңгейге дейін жоғарылатып, аралық тізбекті іске қосу деп есептеуге болады.
Электрондар потенциал айырмашылығына байланысты төмен жылжып, аралық тізбектің соңына дейін жетіп, АТФ синтезін қамтамасыз еткеннен соң, олар НАДФ + -ты тотықсыздандыратындай, теріс потенциалды деңгейге көтерілуі қажет. Дәл осы қызметті I фотожүйе атқарады.
3. Оттегінің бөліну жүйесі. Пигмент Р + 680 электронын ФФ акцепторға беріп, бос катион радикалға айналады. Электрондардың одан әрі тасымалдануы үшін ол тотықсызданып алғашқы қалпына P 680 қайтып келуі керек. Бұған қажетті электрондар судан белінеді. Осы реакцияның нәтижесінде фотосинтездің қосымша өнімі - оттегі бөлінеді. Электрондардың судан Р + 680 пигментке тасымалдану мүмкіндігі олардың потенциал айырмашылығына байланысты. Шамамен Р + 680 потенциалы - +0, 9 В, судыкі - +0, 82 В. Демек, электрондар судан Р + 680 пигментке төмендеп (+0, 82 ден +0, 9 В) ауысады.
Бірақ, электрондардың осылайша тасымалдануын жүзеге асыратын жүйелердің тектері түпкілікті анықталды деп айтуға дәлелдер жеткіліксіз.
Дегенмен, судың тотығып О 2 бөлінуі, су мен Р + 680 аралығында Z деп белгіленетін тотығу-тотықсыздану жүйесінің және Са 2+ , Мп 21 , С1 - иондарының қатысуымен жүзеге асады деген ұғымды көптеген зерттеушілер қолдайды. Шпинат хлоропластарын қараңғылықта 40 минут сақтап, одан соң күшті жарық сәулесімен әсер етгі. Әрбір жеке жарқыл әсерінен бөлінген оттек мөлшері полярограф арқылы анықталды. Бірінші жарқыл әсерінен оттек бөлінбейді, екінші жарқыл әсерінең өте аз мөлшерде бөлінетіндігі байқалды. Оттектің ең көп мөлшерде бөлінуі үшінші жарқылдан кейін байқалады. Оттектің ең көп мөлшерде бөлінуі әр төрт жарқылдан кейін, яғни жетінші, он бірінші ж, т. б. жарқылдарға тура келген. Осы алынған нәтижелер электрон қабылдағыш қараңғылықта тұрақты екі күйде - 1:3 қатынаста (Z және Z 4 ) болады деген қорытынды жасауға мүмкіндік берді. Демек, жарық әсерінен Z біртіндеп Z 4+ күйге айналады. Осы өзгерістер 3-суретте бейнеленген.
Онда Z·F, 80 Q таңбасымен II фотожүйенің реакциялық орталығында бір-бірімен тығыз орналасқан үш бөлік (компоненттер) белгіленген. Екінші фотожүйеге 4 фотон сіңгенде төмен потенциалды b 559 цитохромның 4 молекуласы біртіндеп тотықсызданып Z·P 680 Q жиынтығы Z 4+ P 680 Q күйге айналады. Ол судың екі молекуласымен реакцияланып олардың 4 электронын бөліп алады. Ол электрондар жиналған оң зарядтарды бейтарап реакциялық орталықты бұрынғы ZP 680 Q қалпына қайтарады. Сонымен қатар оттегінің бір молекуласы және 4 протон бөлінеді.
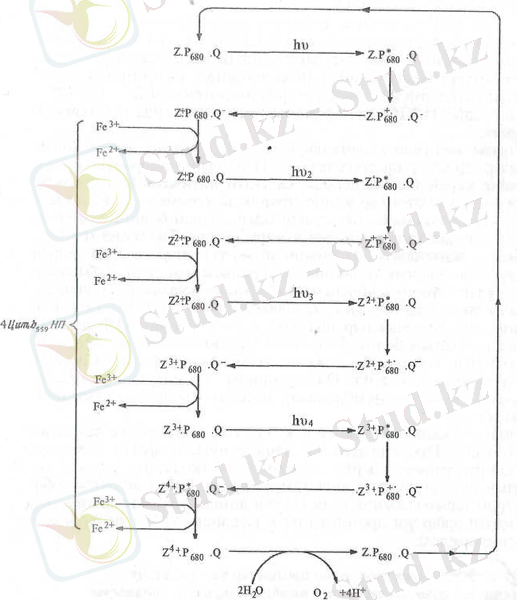
3-сурет. ІІ фотожүйеге байланысты оттегі бөліну жүйесі (Т. Гудвин, Э. Мерсер, 1986) .
Электрондардың циклсіз тасымалдану қатынастары төмендегі тең-деулермен белгіленеді:
4 ФОТОН + Н 2 О → НАДФ Н + 2АТФ + ½ О 2
8 ФОТОН + 2Н 2 О → 2 НАДФ Н + 4АТФ + О 2
Олардан НАДФ + -тың бір молекуласы тотықсызданып, оттегінің бір молекуласы бөліну үшін қанша фотон сіңу қажеттілігін байқауға болады.
1. 3. Электрондардың циклді тасымалдануы
Бұдан бірнеше ондаған жылдар бұрын I фотожүйе айналасында электрондардың циклді тасымалдануын жасанды түрде жүзеге асыруға болатындығы дәлелденген. Ол үшін қосымша тотығу-тотықсыздану жүйелерін сырттан енгізу қажет.
Содан бері бұл процесс хлоропластарда қалыпты жағдайда орын алатындығы анықталды. Бұнда Р* 700 қозған молекулалары электрондарын біртіндеп А 1 , А 2 , А в , ферредоксинге (Фд 590 ), одан b 6 цитохром арқылы пластохинонға береді. Тотықсызданған пластохинон (ПХ-Н 2 ) электрондары f цитохромға, одан пластоцианин және Р + 700 -ге тасымалданады. Бұл тізбекте 2 молекула АТФ синтезделеді. Осы процестің жалпы желісі 4 суретте көрсетілген.
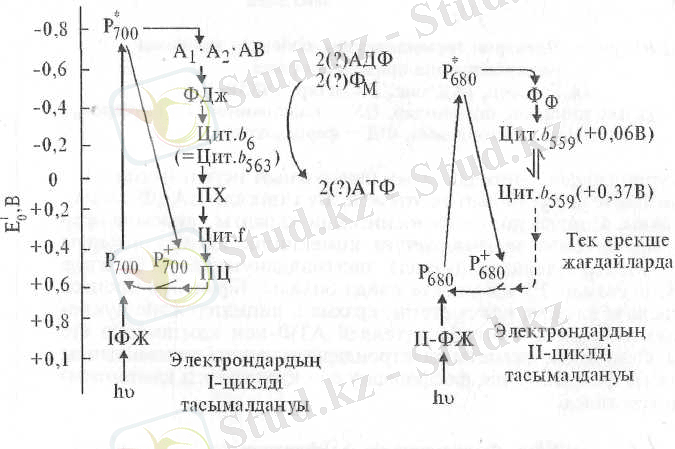
4 сурет. Электрондардың циклді тасымалдану тізбегі (Т. Гудвин, Э. Мерсер, 1986) .
Фотосинтездік I жүйеге байланысты электрондардың циклсіз және циклді тасымалдануын жүзеге асыратын тотығу-тотықсыздану жүйелерін тилакоид мембранасында орналасу болжамы 5-суретте келтірілген. Электрондардың осылайша тасымалдануы I циклді деп аталады. Кейбір жағдайларда өте сирек фотосинтездік II жүйе айналасында байқалатын осы процесс электрондардың II циклді тасымалдануы деп белгіленеді. Бұнда электрондардың тасымалдану жолы Р + 680 пигменттен басталып ФФ және b 559 цитохром арқылы Р + 680 пигментке қайтып тоқтайды. Осы циклдің фотосинтездік фосфорлануға қатысы жоқ деп есептеледі.
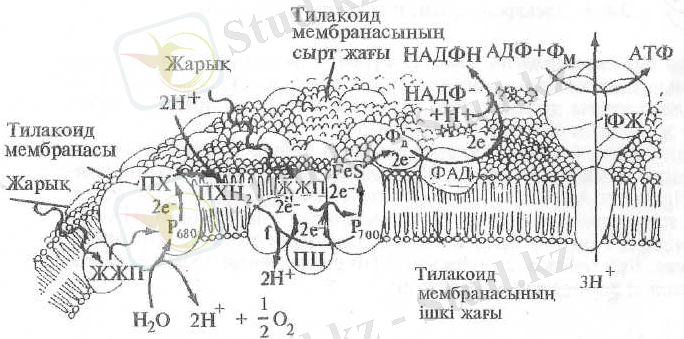
5-сурет. Электрон тасымалдаушы тізбектің тилакоид мембранасында орналасу жүйесі
(А. Гэлстон, П. Дэвис, Р. Сэтгер, 1983) :
ЖЖП - жарық жинаушы пигменттер, ПХ - пластохинон, f - цитохром, ПЦ - пластоцианин, ФД - ферредоксин.
Электрондардың І-циклді тасымалдануының негізгі нәтижесі - АТФ-тың синтезделуі болып есептеледі. Бұл циклде НАДФ тотықсызданбайды. Сондықтан ол фотосинтездің қараңғы сатысына әсер етпейді, тек циклсіз тасымалдануға көмекшілік қызмет атқаруы мүмкін. Электрондардың циклсіз тасымалдануында АТФ және НАДФН, шамамен 2:1 қатынаста пайда болады. Бірақ бұл процесс хлоропластағы энергия қажет ететін, крахмал, липидтер және нуклеин қышқылдарының синтезін түгелдей АТФ-пен қамтамасыз ете алмайды. Осыған байланысты электрондардың циклді тасымалдануы және ондағы фотосинтездік фосфорлану сол қажеттілікті қамтамасыз етеді деп есептеледі.
1. 4 Фотосинтездік фосфорлану
Жоғарыда көрсетілгендей, фотосинтездің жарық сатысында элек-трондардың циклді және циклсіз тасымалдану нәтижесінде бөлінетін энергия АДФ-тың фосфорланып, АТФ-қа айналу процесіне жұмсалады. Бұл процесс тиісінше циклді және циклсіз фосфорлану деп аталады. Фосфорланудың осы екеуінен басқа, псевдоциклді (жалған) деп аталатын түрі де кездеседі. Ол сутегінің тікелей акцепторы - оттегіне байланысты және циклсіз (тікелей) фосфорлануға ұқсас:
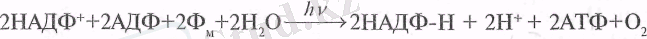
Кейінгі кезде фотосинтездік фосфорлану механизмі ағылшын ғалымы Питер Митчелдің хемиосмостық теориясымен түсіндіріледі.
Бұл теория тилакоид мембранасы арқылы протондардың тасымалдануы туралы ұғымға негізделген.
Электрон тасымалдаушы тізбектің жеке мүшелері тилакоид мембранасында айқас реттеліп орналасқан. Суды тотықтыратын жүйе, пластоцианин, f -цитохромы, сондай-ақ, реакциялық орталық пигменттері мембрананың ішкі жағында, реакциялық орталықтардың акцепторлық бөлігі - флавопротеид (ФД - НАДФ -редуктаза) және НАДФ мембрананың сырт жағында орналасқан (5-сурет) .
Осындағы электрон тасымалдаушылардың біреуі - пластохинон электрондармен қатар протондарды да (Н-иондар) мембрананың сыртынан ішіне қарай тасымалдайды. Осының нәтижесінде тилакоидтың іш жағында протондар концентрациясы көбейеді де, қышқылдығы артады. Тилакоид ішінде судың ыдырауынан бөлінген протондар оның қышқылдығын одан әрі жоғарылатады. Тилакоид мембранасының екі жағындағы рН айырмашылығы энергия көзі болып есептеледі. Осы энергия тилакоид мембранасының сырт жағындағы "саңырауқұлақ тәріздес құрылымның" ерекше өзекшелері (каналдары) арқылы протовдар сыртқа тасымалданғанда пайдаланалуы мүмкін. Бұл өзекшелерде байланыстырушы фактор деп аталатын, АТФ синтезін жүзеге асыра алатын белок-фермент-АТФ-синтетаза бар. Бұл фер-мент АТФ-тың ыдырауын қажетті энергия жеткілікті жағдайда синтезделуін де катализдейді. Ол энергияны "саңырауқұлақ тәріздес құрылымдар" бойымен мембраналар арқылы ағылып жататын протондар жеткізеді.
Басқаша айтқанда, механикалық энергия электрлікке айналған сияқты болады. Протондардың тасымалдануы, олардың тилакоид ішіндегі концентрациясы сыртындағыдан артық болғанша, демек электрондардың ауысуы тоқтағанша ауыса береді. Тізбек арқылы әрбір екі электрон тасымалданғанда тилакоид ішінде 4 протои жиналады. Сыртқа қарай әрбір үш протон қайта ауысқанда, АТФ-тың бір молекуласы синтезделеді.
1. 5 Электрон жене протовдар тасымалдану реакцияларының тилакоид мембраналарында орналасуы
Фотосинтездің фотохимиялық реакцияларына қатысатын белокты ашынтықтар хлоропласт мембраналарында біркелкі орналаспаған.
Фотосинтездік II жүйенің жарық жинаушы пигменттері және пигмент-белоктық жиынтығы негізіиен өзара тығыздалып жанасқан мембраналарда жайғасқан (6-сурет) . Фотосинтездік I жүйе, оның жарық жинаушы жиынтығы көбінесе, ал АТФ-азалық жиынтық (БФ 1 + БФ 0 ) әдетте, мембраналардың өзара нашар жанасқан бөліктерінде орналасады. Цитохром b 6 - f жиынтығы мембраналардың екі түрінде де біркелкі жайғасады. Белокты жиынтықтардың осылайша орналасуы, олардың өзара әрекеттесуі қалайша жүзеге асады деген сұрақ туғызады. Осы әрекеттестілікке себепші болатын факторларға суда ерігіш липофильді пластохинонның оңай жылжығыштығы, пластоцианин ламеллаларының ішкі беті арқылы және ерігіш ферредоксиннің сыртқы беті арқылы ауысатындықтарын жатқызуға болады.
Белокты жиынтықтардың өздері де мембраналарға жанасып жылжи алады. Оған олардың электрлік заряд деңгейі әсерін тигізеді. Мысалы, пластохинонның тотықсыздануы киназаны ырықтандырады және жарық жинаушы пигменттік комплексті фосфорландырады. Фосфорланған ЖЖП-ның теріс заряды көбейіп, оның мембрананың стромалық аймағына жылжуына себепші болады. Оның өзі сіңген жарық энергиясының фотосинтездік I жүйеге ауысуын жоғарылатады. Фотосинтездік жүйенің фотохимиялық ырықтығының артуы пластохинонның тотығуын күшейтіп, киназаны ырықсыздандырады. Фосфатаза болса ЖЖП-ны фосфорсыздандырады. Сонымен ЖЖП-ның қайтымды фосфорлануы фотожүйелер ырықтығының өзара реттеушілік жүйесіндегі кері байланыстылық түзағы болып есептеледі.
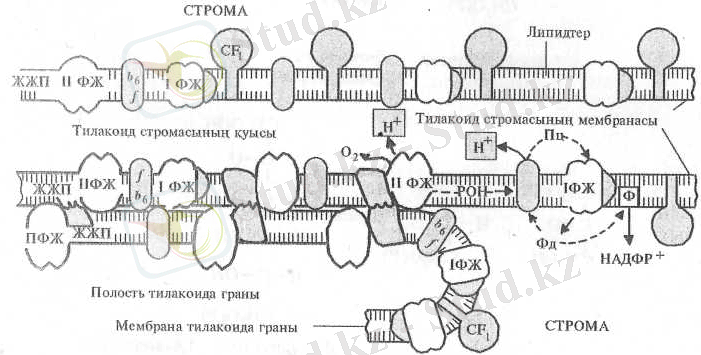
6-сурет. Тилакоид мембраналарында фотосинтездің жарық сатысына қатысатын құрылымдардың орналасуы (В. В. Полевой, 1989) :
ЖЖП - жарық жинаушы пигментттер, 1, 11, ФЖ - фотожүйелер, b - цитохром b, f - цитохром f, БФ - байланыстырушы фактор, ФД - ферредоксин, ПЦ - пластоцианин, ПХ - пластохинон, Р - редуктаза.
2. Фотосинтездің қараңғы сатысы
2. 1 Фотосинтездің С 3 - жолы (Кальвин циіслі)
Кальвин циклінің алғашқы реакциясы сырттан енген СО 2 -нің Д-рибулозо-1, 5-дифосфатпен реакциялануынан басталады; нәтижесінде 3-фосфоглицерин қышқылының (3-ФГК, ) екі молекуласы пайда болады:
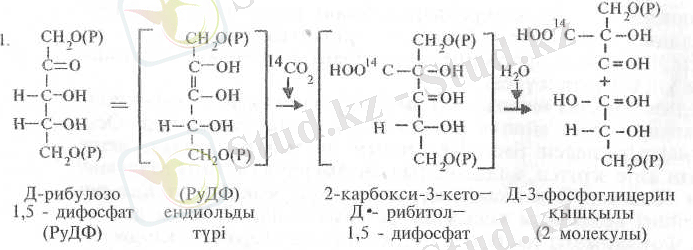
Бұл реакцияны рибулозо-1, 5-дифосфаткарбоксилаза (РуДФ-кар-боксилаза) ферменті катализдейді. Реакцияда көрсетілгендей, рибу-лозодифосфат алдымен таутомерленіп, енолдық күйге ауысады, содан соң көміртегінің екінші атомы арқылы карбоксилденеді.
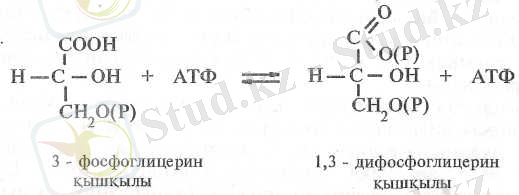
Пайда болған 3-ФГҚ фосфорланып 1, 3-дифосфоглицерин қышқылына (1, 3-ДФГҚ) айналады. Бұл фосфорлану фотосинтездің жарық сатысында синтезделген АТФ-тың есесінен жүзеге асады. Осы реакцияны фосфоглицераткиназа ферменті катализдейді:
Одан соң, 1, 3-ДФГҚ фотосинтездің жарық сатысының өнімі НАДФН-тың қатысуымен тотықсызданып 3-фосфоглицеральдегидке айналады. Реакцияны цитоплазмалық, НАД- пен байланысқан глицеральдегид-3-фосфат дегидрогеназа ферменті катализдейді:
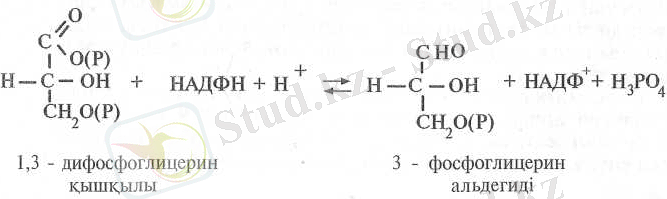
Фосфоглицеральдегид молекулалары Кальвин циклінде төрт түрлі бағытта өзгерістерге ұшырайды. Олардың біріншісі төмендегі реак-циядағыдай 3-ФГА изомерленіп (ДГАФ) айналуы. Бұл реакция триозофосфатизомераза ферментінің қатысуымен жүзеге асады:

3-ФГА-тің өзгеруінің екінші жолында ол ДГАФ молекуласымен қосылып, ферментінің әсерінің Д-фруктозо-1, 6-дифосфат (Ф1, 6ДФ) молекуласын түзеді:
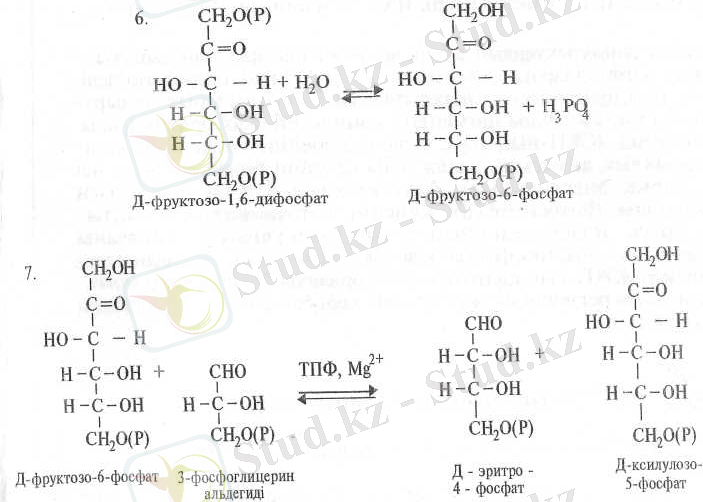
Пайда болған Ф-1, 6-ДФ фруктозодифосфатаза ферментінің әсерінен фосфорсызданып Д-фруктозо-6-фосфатқа (Ф-6-Ф) айналады (6) .
3-ФГА пайдаланылуының үшінші жолында ол фруктозо-6-фосфат молекуласымен реакцияланып Д-ксилулозо-5-фосфат және Д-эритрозо-4-фосфат (КСУ-5-Ф), (Э-4-Ф) молекуларын түзеді (7) .
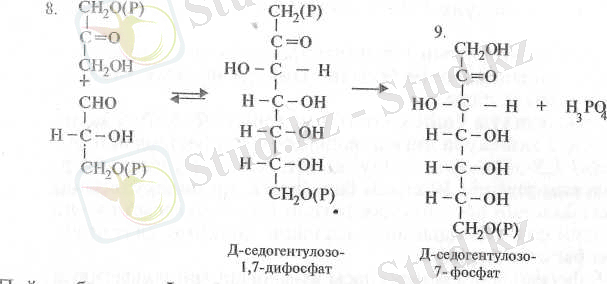
Пайда болған эритрозо-4-фосфат ре-акцияласып седогептулозо-1, 7-дифосфатқа айналады (8) . Бұл реакцияға ферменті қатысады.
Осыдан соң седопгептулозо-1, 7-дифосфат дефосфорланып седо-гептулозо-7-фосфатқа айналады (9) . Бұл реакция седогептулозоди-фосфатазамен катализденеді. Келесі 3-ФГА-нің өзгеріске түсуінің 4 жолывда, транскетолаза ферментінің қатысуымен, ол седогептулоза молекуласымен реакцияласады да нөтижесінде ксилулозо-5-фосфат және рибулозо-5-фосфат молекулалары түзіледі (10) :
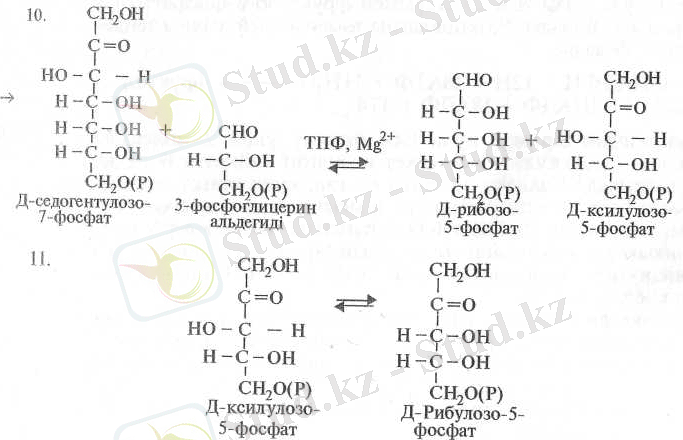
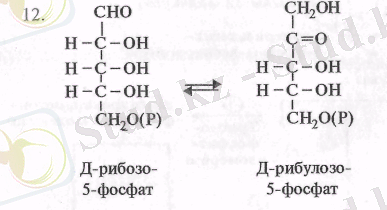
Одан әрі ксилулозо-5-фосфат рибулозофосфат-3-эпимераза ферментінің әсерінен көміртегінің үшінші атомынан эпимерленіп Д-рибулозо-5-фосфатқа айналады (11) . Рибозо-5-фосфаттың рибозо-фосфатизомераза ферментінің өсерінен изомерленуінеи рибулозо-5-фосфат пайда болады (12) .
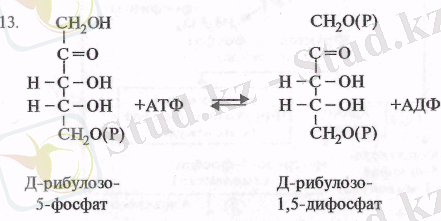
Ең соңында, пайда болған рибулозо-5-фосфат фотосинтездік фос-форлану нәтижесінде синтезделген АТФ-тың есесінен, фосфорибулокиназа ферментінің қатысуымен рибулозо-1, 5-дифосфатқа айналады (13) :
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz