Өсімдіктердегі тыныс алудың даму тарихы, химиялық негіздері және ферменттік жүйелері

Мазмұны
Кіріспе. . 3
I. Тыныс алу туралы ілімнің даму тарихы
1. 1. Ең алғашқы деректер 4
1. 2. Оттегінің ырықтануы туралы ұғымдар. 5
1. 3. Сутегінің ырықтануы туралы ұғымдар. . 6
1. 4. Тыныс алу мен ашу процестерінің байланыстылығы. 9
II. Тыныс алудың ферменттік жүйелері.
2. 1. Оксидоредуктазалар 12
ІІІ. Тыныс алу заттарының (субстрат) негізгі тотығу жолдары.
3. 1. Гликолиз (Тыныс алудың анаэробты сатысы) . . 21
3. 2 Гликолиз өнімдерінің анаэробты өзгеру жолдары. Ашу процестері . 24
3. 3. Ди- және трикорбон қышқылдар циклі . 27
3. 4. Глиоксилаттық цикл 32
VІ. Қорытынды. 34
V . Қолданылған әдебиеттер тізімі 35
Кіріспе
Тыныс алу жер бетіндегі барлық организмдерге, кез келген органға, жеке ұлпаларға және клеткаларға тән өте күрделі процесс. Организмнің тыныс алуы тоқтаса тіршілігі де тоқтайды.
Өсімдіктердің тыныс алуында фотосинтез нәтижесінде пайда болған өнімдер оттегінің әсерінен тотығып ыдырап, қарапайымдау аралық заттарға және ең соңында анорганикалық өнімдерге (С02 және Н20) айналады. Органикалық күрделі қосындылардың біртіндегі ыдырауына байланысты бөлінген энергия клетканың бөлінуіне, өсу, даму және көбею, судың және қоректік заттардың сіңіп, таралу, осыларға байланысты алуан-түрлі синтездік процестерге жұмсалады. Сонымен қатар сатылылықпен пайда болатын аралық қосындылар өсімдік организміндегі көптеген зат алмасу процестерінің бастамасы бола алады. Жалпы айтқаңда тыныс алу, біріншіден организм тіршілігі үшін қажет энергияның ішкі көзі, екіншіден негізгі зат алмасу процестерін өзара байланыстырушы орталық болып есептеледі.
Өсімдіктің тыныс алуына, негізінен, көмірсулар пайдаланады. Бірақ өсімдіктің түрлеріне, өсіп-даму кезендеріне, сыртқы ортаның жағдайларына байланысты басқа да органикалық қосындылар (май, белок) пайдаланылуы мүмкін.
Тыныс алудағы химиялық өзгерістердің жалпы теңдеуін төмендегіше бейнелеуге болады:
С6Н206 + 602 = 6С02 + 6Н20 + 2867 кДж
Яғни, глюкозаның бір молі (180 г) тотыққанда оттегінің 6 молі сіңіп, көмірқышқыл газының 6 молі, судың 6 молекуласы және 2867 кДж энергия бөлінеді. Бұл теңдеуден тыныс алу фотосинтезге карама-қарсы процесс екендігін байкауга болады.
І. Тыныс алу туралы ілімнің даму тарихы
1. . Ең алғашқы деректер.
Тыныс алу туралы ұғымдар өсімдіктер физиологиясы ғылымның жеке саласы ретінде қалыптаса бастаған кезден-ақ пайда болды. Бірақ, ол туралы ілімнің дамуы XVIII ғасырдың соңғы ширегінен басталды. Бұл кезде Дж. Пристли, А. Л. Лавуазье және басқа ғалымдардың зерттеулері нәтижесінде ауа құрамы анықталып, оттегі (1774 ж. ) ашылған болатын. Хайуанаттардағы тыныс алу процесін және жануды қатар зерттеп А. Л. Лавуазье 1773-1783 ж. ж. тыныс алуда және жануда оттегі сіңіп, көмірқышқыл газы бөлінеді деген тыжырым жасады. Ол өзінің тәжірибелеріне сүйеніп жану процесіне оттегі қажет, ал тыныс алу - тірі организмде коректік заттардың баяу жануы деп ұйғарды.
Я. Ингенхауз 1778-1780 ж. ж. жасыл өсімдік жарық жағдайына байланысты С0 2 сіңіріп, О, бөлумен қатар, 0 2 сіңіріп, СО 2 бөліп шығаратындығын дәлелдеді. Шамамен, 20-25 жьілдан соң Н. Т. Т. Соссюр өз тәжірибелерінің нәтижесінде жасыл өсімдіктерде газ алмасудың өзара қарама-қайшы екі түрі бар екендігін дәлелдеді. Ол өсімдіктің жасыл бөліктерінен С0 2 бөлінуі және оттегінің сіңуі тек қараңғылықта ғана, ал түссіз бөліктерінде - түнде де, күндіз де байқалатындығын көрсетті. Өте дәл әдістермен жүргізілген осы зерттеулер нәтижесінде қалыптасқан қорытындылар коп уақытка дейін ғалымдар арасында қабылданбады. Көптеген беделді зерттеушілер, өсімдік организмінде тыныс алуға бейімді дербес органның жоқтығына сүйеніп, осы процестің болатындығына күмәнденді. Сондай-ақ бір организмде қатарынан қарама-қарсы екі процесс - фотосинтез және тыныс алу болуы мүмкін емес деп ұйғарылды.
Осыған байланысты 1842 ж. атақты неміс ғалымы Ю. Либих өсімдіктерде тыныс алуды жоққа шығарып, түнде С0 2 -нің фотосинтезге пайдаланылмаған қалдығы бөлінеді деген тұжырым жасады.
Бірақ XIX ғасырдың соңғы, XX ғасырдың алғашқы кезендеріндегі зерттеулердің нәтижесінде хайуанаттар мен өсімдіктердегі тыныс алу біртектес екендігі туралы жаңа мәліметтер алынды. Тыныс алу процесіне пайдаланылатын заттардың ең негізгісі көмірсулар болатындығы дәлелденді. Осы ілімнің жан-жақты терендеп дамуына үлес қосқан ғалымдар И. П. Бородин, Х. Ф. Шейнбайн, А. Н. Бах, В. И. Палладии, С. П. Костычев, О. Варбург, Х. Виланд зерттеулері биологиялық жағдайда тірі клетканың сырттан энергия пайдаланбай органикалық заттарды тотықтыру қабілеттілігін қамтамасыз ететін факторлардың тегін табуға бағытталды.
XX ғасырдың 20-шы жылдарынан бастап жүргізілген Д. Кейлин, Г. Эмден, О. Мейергоф, Х. Кребс, А. Сент-Дьерди, В. Христиан, Д. Грин, С. Очоа, Х. Теорель, Д. М. Михлин және т. б. ғалымдардың зерттеулерінің нәтижесінде тыныс алу процесінің химиялық негіздері мен ферменттік механизмдері анықталды.
Алдымен хайуанаттар тыныс алуындағы ашылган зандылықтар өсімдіктерге де тән екендігі, сонымен қатар соңғыларда осы процеске байланысты катализдік жүйелер, хайуанаттардағыға қарағанда, күрделірек болатындығы анықталды.
Қазіргі кездегі көзқарастарға сәйкес оттекті (аэробтық) тыныс алу жерге жақын атмосферада бос оттегі пайда болған соң ғана қалыптасты. Жасыл өсімдіктер жоқ кезде біздің планетадағы организмдерде энергияның алмасуы анаэробтық жолмен, яғни оттегінін қатысуынсыз жүзеге асқан.
Анаэробтық процестің аэробтық түрге ауысуы тыныс алуға жұмса-латын заттар молекуласындағы энергияның толығырақ және тиімді пайдаланылуына мүмкіндік туғызды. Осы өзгерістің өзі, эволюция процесіңде клеткадағы энергиясы мол күрделі қосындыларды біртіндеп энергиясы аз күйге дейін ыдырататын, жаңа ферменттік жүйелердің қалыптасуына байланысты болды.
Аэробтық процестің қалыптасуына байланысты анаэробтық процесс түгелінен жойылып кеткен жоқ. Керісінше, ол тыныс алудың белгілі бір сатысы болып қалды.
1. 2. Оттегінің ырықтануы туралы ұғымдар
Оттегі молекуласы - О химиялық жағынан реакциялық қабілеттілігі төмен, өздігінен биологиялық ортада (организмде) тыныс алуға жұмсалатын заттарды тотықтыра алмайды. Бұндай реакцияларға қатысу үшін ол алдымен ырықтануы (активтенуі) қажет.
Өзінің тотығу процестерінің химиялық негіздеріне арналған зерттеулерінде А. Н. Бах молекулалық оттегі тотықтырғыш болуы үшін оның қос байланысы үзіліп босануы қажет деген тұжырымға келді. Ол өзінің осы пікірін 1897 ж. "биологиялық тотығудың асқын тотықтық теориясы" түрінде қалыптастырды.
Осы теорияға сәйкес оңай тотыққыш қосынды - А оттегімен әрекеттесіп қос байланысы үзіліп, асқын тотыққа - А0 2 айналады. Осы қосынды В қосындысымен әрекеттесіп оны тотықтырады. Содаң соң осы реакция оттегінің екінші атомымен және В қосындысының екінші молекуласымен қайталанады. Нәтижесінде алғашқы қосынды - оттегінің акцепторы - А толығынан тотықсызданып, В қосындысы толығымен тотығады:
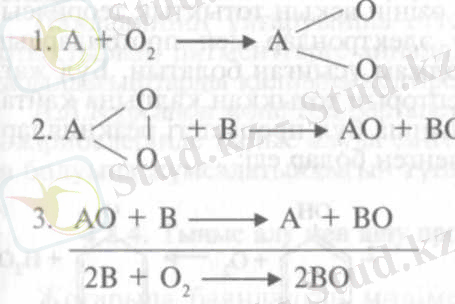
А. Н. Бахтың ұйғаруынша екінші және үшінші реакцияларға пе-роксидаза қатысады. Оттегін активтендіретін А қосындысын фермент деп ұйғарып, А. Н. Бах оны "оксигеназа" деп атады. Кейінірек, ол өсімдік ұлпалары жарақаттанғанда ауаның әсерінен қоңырланатын қосындылар осы заттар деп қорытынды жасады. Дәл осындай оңай тотығатын заттар ғана оттегін байланыстырып асқын тотыққа айналады.
Осыдан көп кейінірек, яғни 1955 ж. жапония ғалымы О. Хаяиши, америка ғалымы Г. С. Мэзон өздерінің қызметтестерімен бірге жаңа әдістерді қолданып, оттегінің органикалық қосындыларға қосылу мүмкіндіктерін зерттеді. Хаяиши оттегінің ауыр изотопын қолданып, пирокатехиннің пирокатехаза ферментінің әсерінен тотығу процесін зерттеді.
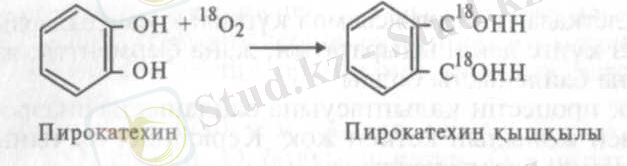
Бұл реакциядан пирокатехин қышқылындағы оттегі таңбалы екеңдігі байқалып тұр. Сонымен өте дәл тәжірибенің нәтижесінде оттегінің молекуласы органикалық қосындылардың құрамына қосыла алатындығы дәлелденді.
Қазіргі кезде, биологиялық тотығудың асқын тотықтық теориясына байланысты, оттегінің органикалық қосындыларға ену жолдарының тыныс алуға қатысты емес екендігі белгілі болды. Неміс биохимигі О. Г. Варбург 1921 ж. тыныс алуға басытқылардың әсерін зерттеп, көміртек тотығып және синиль қышқылы оттегінің сіңуін тежейтіндігін байқады. Ағылшын биохимигі Д. Кейлин 1925 ж. клеткаларда оттегінің сіңуін жеделдететін цитохромоксидаза болатындығын нақты дәлелдеп, басқа да цитохромдарды ашты. Содаң соң барлық аэробты организмдерде цитохромдар табылып, тыныс алудың ең соңғы сатысы оттегіне электрондар мен протондардың тасымалдануынан судың пайда болуымен аяқталатындығы анықталады.
1. 3. Сутегінің ырықтануы туралы ұғымдар
Органикалық қосындылар басқа жолмен де, яғни сутегінің бөлінуі арқылы да тотығады. Кезінде Бах өзінің асқын тотықтық теориясымен қатар, биологиялық тотығу электрондар мен протондардың бөлінуіне де байланысты деген болжам ұсынған болатын. Бұл жағдайда оттегі сутегінің бірінші акцепторын тотыққан қалпына қайтару қызметін атқарады. Бахтың ойынша бұл процестегі реакциялардың бір ізділігі төмендегіше белгіленген болар еді:
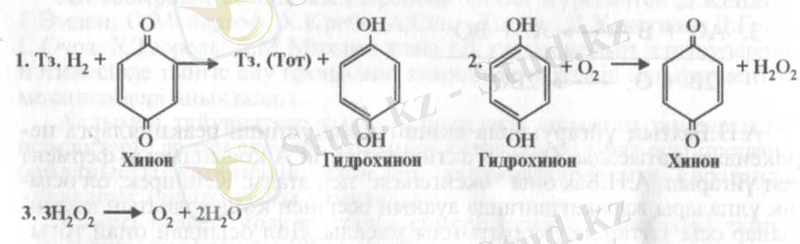
Бахтың бұл болжамын В. И. Палладии тыныс алу химизмінің нақты теориясына дейін дамытты. Өзі 1903-1916 ж. ж. тыныс алу пигменттерін зерттеумен айналысып, химиялық реакцияларды талдауға өте ыңғайлы жасанды үлгі ашты. Ол кәдімгі метилен көгі болатын. Егер, ішіне метилен көгінің ерітіндісі кұйылған пробиркаға бидайдың өнген ұрығының бірнешеуін салып, ауаны (оттегін) ығысғырып, тығындап қойса, біраз уақыттан кейін ерітінді түссізденеді. Одан соң пробирканың аузын ашып ішіндегі ерітіндіні шайқаса (ауа енгізсе) ол қайтадан көгереді. Сонымен, оттегімен әрекеттескенде бояғыш тотығып көгереді де, тірі тұқым өскіні бояғышты тотықсыздандырады. Бұндағы метилен көгінің тотығуы оттегінің қосылуынан емес, сутегінің бөлінуіне байланысты. Бояғыш электрондар мен протондардың енуіне байланысты тотықсызданады.
Өзінің осы және басқа тәжірибелерінің нәтижесінде Палладии 1912 ж. тыныс алу химизмінің жалпы теориясын қалыптастырып, оның негізгі теңдеуін анаэробты (1) және аэробты (2) деп бөлді:
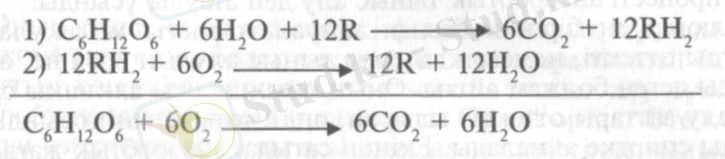
мұндағы, R - боялған тыныс алу пигменті, тыныс алуға жұмсалатын заттан (субстрат) сутегін бөліп алады, RH 2 - түссіз тыныс алу хромогені.
Келтірілген тендеулерден көрінгендей, тыныс алудың бірінші, анаэробтық сатысында су және ерекше тыныс алу хромогендері қатысып, глюкоза сутегінің бөлінуінен тотығады. Екінші - аэробтық реакцияда хромогендер қайтадан тотығып, одан бөлінген сутегі отгегіне қосылып, су пайда болады. Бұл жерде судың екі түрлі маңызын -бір жагынан сутегінің, екіншіден оттегінің көзі екендігін байқауға болады.
Палладиннің ұйғаруынша оттегінің молекуласы хромогендерді тотықтырып, пигменттерге айналдыру ролін атқарады, яғни сутегін қабылдағыштарды қалпына келтіреді.
А. Л. Курсанов және Б. Б. Вартапетян 1955 ж. 18 0 қолданып жүргізген тәжірибелерінде тыныс алуда сіңген оттегі шындығында судың пайда болуына жұмсалатындығын түпкілікті дәлелдеді.
1. 4. Тыныс алу мен ашу пронес терінің байланыстылығы
Жоғарыда баяндалған мәліметтерден тыныс алу процесі екі негізгі сатыдан тұратындығы айқын көрінеді. Бірінші, анаэробтық сатыда тыныс алуға жұмсалатын зат (көмірсулар) пирожүзім қышқылына, екінші - аэробтық жағдайда соңғы өнімдерге - С0 2 және Н 2 0 дейін өзгереді. Сонымен тыныс алу мен ашу процестерінің арасында белгілі байланыстылық байқалады. Осы процестердің арасындағы генетикалық байланыстылық туралы ілімді С. П. Костычев қалыптастырды.
Дегенмен, осы бағытта ең алғаш зерттеулер жүргізген ғалымдардың бірі - Н. Соссюр XIX ғасырдың бас кезінде-ақ жасыл өсімдіктер оттексіз ортада да СО 2 бөліп шығарады деген болатын.
Сол шамаларда Л. Пастер жасыл өсімдік ұлпалары анаэробтық жағдайда көмірқышқыл газын бөлумен қатар спиртті, яғни спирттік ашу процесінде пайда болатын басқа қосындыларды да бойыңод жинай алады деп ұйғарды. Ол жасыл өсімдіктен бөлінетін С0 2 , ешқандай микроорганизмдердің қатысуынсыз, тікелей өсімдіктің өз ұлпаларындағы реакциялардың нәтижесі екендігін дәледдеді.
Осы кездері неміс физиологы Е. Пфлюгер хайуанат (көлбақа) оттексіз ортада біраз уақытқа дейін тіршілігін сақтай алатындығын байқады. Осы жағдайда хайуанаттан бөлініп шығатын көмір-қышқылын Пфлюгер органикалық молекулалардағы байланысқан оттегінің әсерінен ыдыраудың өнімі деп қарады. Сондықтан осы процесті ол молекулалық тыныс алу деп атады. Кейінірек С. П. Костычев бұл процесті анаэробтық тыныс алу деп атауды ұсынды.
Е. Пфлюгер ең бірінші болып хайуанаттардағы молекулалық тыныс алуды оттекті, немесе қалыпты тыныс алудың ерекше, алғашқы сатысы деген болжам айтты. Оның ойынша осы алғашқы сатыда тыныс алу заттары оттексіз ыдырап, оңай тотығатын қосындыларға, мысалы спиртке айналады. Екінші сатыда, анаэробтық жағдайда пайда болған спирт ауадағы оттегінің әсерінен соңғы өнімдерге - С0 2 және суға дейін ыдырайды. Осы көзқарасты XIX ғасырдың аяқ кезіндегі көрнекті физиолог-ботаниктердің бірі В. Пфеффер де жақтады. Өз зерттеулерінің нәтижесіне сүйеніп, Пфеффер мен Пфлюгер тыныс алу механизмін сипаттайтын төмендегідей екі теңдеуін ұсынды:
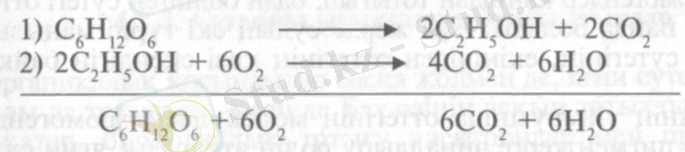
Сонымен қатар Пфеффер спирт өсімдіктердің қалыпты тыныс алуының аралық өнімі және тыныс алу ашу процесімен байланысты деген көзқарасты да ұсынды.
Тыныс алу мен ашу процестерінің өзара байланыстылыгы туралы дөлелдер В. И. Палладии мен С. П. Костычев тәжірибелерінде де алынған болатын. Олар өнген тұқымдарда және өркендердің осу нүктелерінде анаэробтық және аэробтық жағдайларда да спирт пайда болатындығын байқады.
Осы алынған деректер, өте маңызды болғанымен, калыпты тыныс алу және ашу процестері арасындағы байланыстылық негізін толық ашуға мүмкіндік бере алмады. Сондай-ақ оттексіз жагдайда өсімдіктердің тыныс алу және ашытқылардағы спирттік ашу процестерінің ұқсастығы туралы сұрақ та шешусіз қалды.
Осы сүрақтарды шешуде С. П. Костычев зерттеулері үлкен маңызға ие болды. Ол өсімдіктің қалыпты - аэробтық тыныс алуында спирт төмендегідей екі себептен аралық зат бола алмайтындығын тәжірибе жүзінде дәлелдеді: біріншіден, спирт улы зат, өсімдік ұлпасында көп мөлшерде жиналмайды, екіншіден, ол өсімдік ұлпаларында глюкозаға қараганда өте нашар тотығады. Бұл дәлел Пфлюгер мен Пфеффердің көзқарастарына карама-қайшылық туғызады.
Өз зерттеулерінің нәтижелеріне сүйене отырып С. П. Костычев тыныс алу мен ашудың генетикалык байланыстылыгы туралы теориясын дамытты:

Оның теориясына сәйкес екі физиологиялық құбылыс - тыныс алу және ашу өзара бөлек емес, керісінше өзара тікелей байланысты. Қанттың өзгеруінің алғашқы сатысы - тыныс алу субстратының анаэробтық ыдырауы екі процеске де бірдей ортақ. Өте көп сатылы бір ізді өзгерістер арқылы осы ыдырау нәтижесінде тыныс алуға да, ашуға да бірдей ортақ аралақ қосындылар пайда болады. Осы екі процестің бағыттары, қанттың ыдырауы белгілі сатыга жеткен кезде ғана ажырайды.
Қазіргі кезде Костычевтің, неміс биохимигі К. Нейбергтің және басқалардың зерттеулеріне негізделген көзқарастарға сәйкес тыныс алу және ашу процестерінің барлық түрлері өзара пирожүзім қышқылы арқылы (ПЖҚ) байланысады:
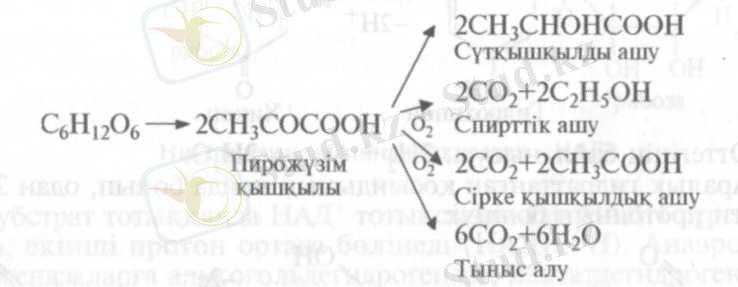
Қорытып айтқанда Костычевтің тыныс алу мен ашудың генетикалык байланыстылығы туралы теориясы толығымен дәлелденді.
ІІ. Тыныс алудың ферменттік жүйелері
Тірі ұлпалардағы органикалық қосындылардың тотығуы сутегін және молекулалық оттегін ырықтандыратын, аралық сутегін тасымалдаушылық қызметін атқаратын алуан түрлі ферменттердің қатысу ымен жүзеге асады.
Тыныс алудың ферменттік жүйелері, химиялық механизмдері сыртқы орта жағдайларының үздіксіз өзгергіштігіне бейімделіп қалыптасқан.
Өсімдіктердегі тотығу-тотықсыздану аппаратының құрылымын сипаттайтын үш түрлі ерекшелігін атауға болады. Біріншіден, өсімдікте тотықтыру жүйелерінің алуан түрлі болуы. Осыған байланысты клеткадағы белгілі бір реакцияға, немесе функциясы жағынан тектес реакцияларға бір ғана емес, бірнеше фермент қатысуы мүмкін. Бұның өзі өсімдіктегі биологиялық тотығу жолдарының, оған катализдік жүйелердін қатысу үлесінің құбылмалылығын қамтамасыз етеді.
Тотығу процесінің механизмдері мен жолдарының құбылмалылығы өсімдіктердің сыртқы орта жағдайларына бейімделушілік мүмкіндіктерін молайтады.
Тотығу аппаратының екінші ерекшелігіне катализдік жүйелердің атқаратын қызметінің көптігін, яғни катализдеушілердің көп қасиеттілігін жатқызуға болады.
Үшіншіден, тотығу аппаратының орналасу орындары да көп болады. Қазіргі кезде протопластың құрылымды бөліктерінің барлығында дерлік өздеріне тән тотығу жүйелерінің болатындығы анықталды.
Электроңцардың бөлінуіне байланысты тотығуды төрт түрлі жолы
бар:
1. Электрондардың тікелей ауысуы: Ғе 2+ Ғе 3+ .
2. Сутегінің бөлінуі:
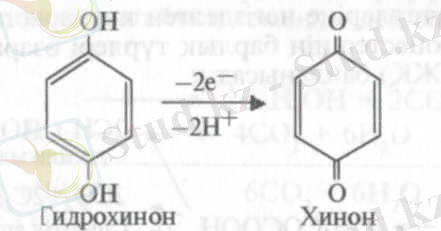
3. Оттегінің байланысуы: 2Н 2 + 0 2 -» 2Н 2 0.
4. Аралық гидраттанған қосындының пайда болып, одан 2 электрон мен протонның бөлінуі:
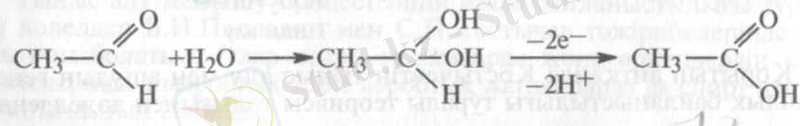
2. 1. Оксидоредуктазалар
Бір заттың (электрон мен протон беруші) тотығуы екінші заттың (оларды қабылдаушы) тотықсыздануымен қатар жүретіндіктен, осы реакцияларды катализдейтін ферменттер оксиредуктазалар деп аталады. Олардың барлығы да ферменттердің І-класына жатады.
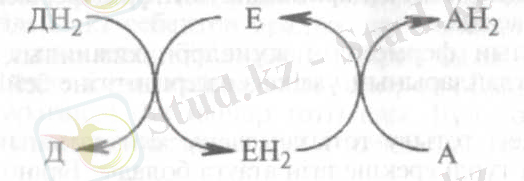
Донор (Д) - электрон мен протонды беруші, акцептор (А) - оларды қабылдаушы, энзим (Е) ауысу реакциясын іске асырады.
Оксидоредуктазалар үш топқа бөлінеді:
а) анаэробты дегидрогеназалар электрондарды және протондарды оттегіне емес, оған дейінгі аралық қабылдағыштарға ауыстырады;
б) аэробты дегидрогеназалар электрондарды және протондарды әр түрлі қабылдағыштарға, соның ішінде оттегіне ауыстырады;
в) оксидазалар электрондарды тек оттегіне жеткізеді.
Анаэробты дегидрогеназалар. Бұл екі бөлікті, коферменті НАД + () болатын ферменттер:
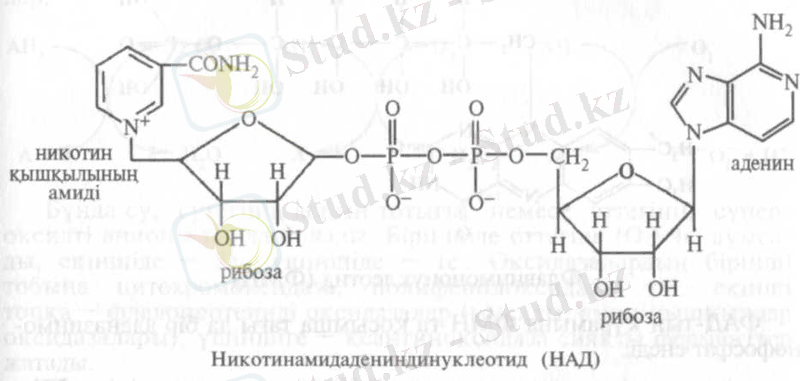
Субстрат тотыққанда НАД + тотықсызданған НАДН түрге ауысады да, екінші протон ортаға бөлінеді (НАДН+Н) . Анаэробты дегидрогеназаларға , лактатдегидрогеназа, малатдегидрогеназа, т. б. жатады. Коферменті, НАД-тан бір фосфаты артық, НАДФ болатын () анаэробты ферменттер де бар: , глюкоза-6-фосфатдегидрогеназа, 6-фосфоглюконат-дегидрогеназа және т. б.
Ферменттік заттың (субстраттық) ерекшелігі оның белокты бөлігіне байланысты. Көптеген НАД- және НАДФ-тәуелді дегидрогеназалар екі валентті металдар иондарының болуын қажет етеді. Мысалы, құрамында мырыш иондары бар.
Анаэробты дегидрогеназалардың тотыққан және тотықсызданған түрлері НАД(Ф) -трансгидрогеназа ферментімен катализденетін реакцияда бір-біріне ауыса алады:

Анаэробты дегидрогеназалар сутегі, немесе электрон мен протонды әртүрлі аралық тасымалдаушыларға және аэробты дегидрогеназаларға ауыстырады.
Аэробты дегидрогеназалар. Бұлар да екі бөліктен тұратын флавинді (флавопротеиндер) деп аталатын ферменттер. Олардың құрамына белоктан басқа, солармен тығыз байланысқан топ - рибофлавин (витамин В 2 ) енеді.
Бұл топта коферменттің екі түрі: флавинмононуклеотид (ФМН) немесе Варбургтың сары тыныс алу ферменті және (ФАД) кездеседі.
ФМН (рибофлавин-5-фосфат) құрамына гетероциклді азотты негіз - диметилизоаллоксазин, рибит спирті (рибоза туындысы) және фосфат енеді:
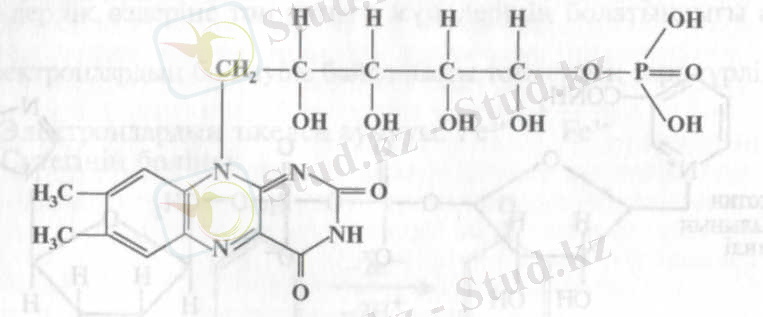
Флавинмононуклеотид (ФМН)
ФАД-тың құрамына ФМН-ға қосымша тағы да бір аденазинмо-нофосфат енеді:
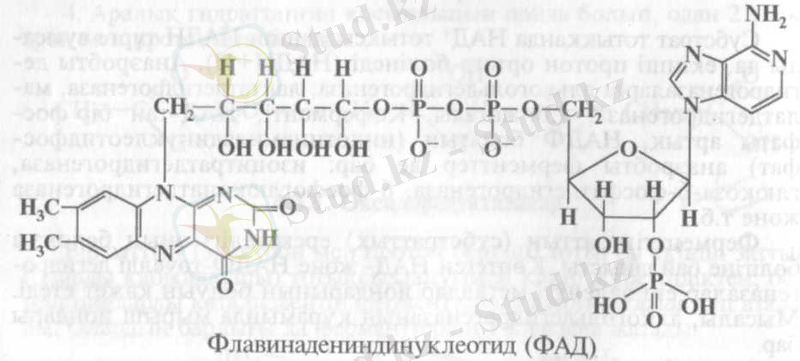
ФМН мен ФАД арасында электрондар мен протондар ауысуы реакциясында ырықты топ қызметін изоаллоксазин атқарады. То-тықсызданған тасушымен, мысалы НАДН, әрекеттесу төмендегіше жүреді:
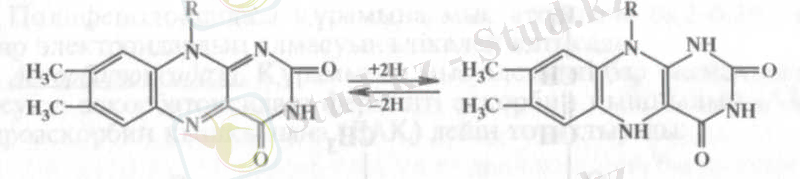
Құрамына ФАД енетін дегидрогеназаның мысалы ретінде сукци-натдегидрогеназаны келтіруге болады. Аэробты дегидрогеназаларға электрондар мен протондарды беретін (донор) - анаэробты дегидрогеназалар, кабылдайтындар (акцептор) - хинон, цитохромдар жөне оттегі.
Оксидазалар. Бұлар электрондарды тотығатын заттан алып, тек оттегіне ғана тасушы (жеткізуші) аэробты дегидрогеназалар:
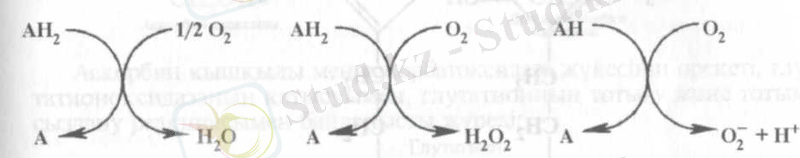
Бұнда су, сутегінің асқын тотығы, немесе оттегінің супероксидті анионы пайда болады. Біріншіде оттегіне (0 2 ) 4е - ауысады, екіншіде - 2е - , үшіншіде - 1е - . Оксидазалардың бірінші тобына цитохромоксидаза, полифенолоксидаза, т. б. екінші топқа - флавопротеинді оксидазалар (мысалы, амин қышқылдар оксидазалары), үшіншіге - ксантиноксидаза сиякты ферменттер жатады.
Сутегінің асқын тотығы (Н 2 0 2 ) және оттегінің супероксидті анионы (0 2 ~) өте уытты, сондықтан клеткада арнайы ферменттердің әсерінен Н 2 0 және 0 2 - -ге өзгереді.

Цитохромдық жүйе. Оксидазалардың ішінде өте маңызды рөл атқаратын темірлі ферменттер және цитохромдық жүйеге жататын тасымалдаушылар болып есептеледі. Оған цитохромдар және цитохромоксидаза енеді. Белгілі бір ізділікпен электрондар тасымалдану процесіне енген олар электрондарды флавопротеиндерден молекулалық оттегіне жеткізеді. Цитохромдық жүйенің барлық мүшелерінің құрамында темірпорфиринді белоксыз топ болады:
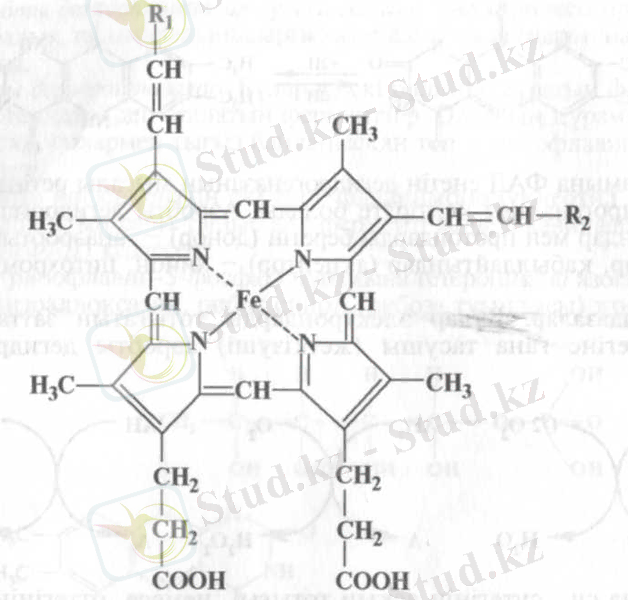
Цитохромдар арқылы электрондар тасымалданғанда темір қайтымды тотығу-тотықсыздану байланысты валенттілігін өзгертеді. Тыныс алу тізбегіндегі электрондардың тасымалдану бағыты цитохромдардың тотығу-тотықсыздану деңгейіне (потенциалына) байланысты анықталады:
цит. цит. цит. цит.
b ►с 1 ►с ► аа 3 ► 0 2
Бұл жүйенің ішінде тек цитохромоксидаза ғана электрондарды тікелей оттегіне жеткізе алады. басытқыларына СО, цианид, азид жатады. Өсімдік митохондрияларында одан басқа, цианидпен басытқыланбайтын және альтернативті оксидаза деп аталатын фермент бар.
Өсімдік клеткаларында митохондриядан тыс орналасатын поли-фенолоксидаза, аскарбатоксидаза, т. б. оттегімен әрекеттесе алатын ферменттер кездеседі.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz