Өсімдік генетикасы мен селекциясының молекулалық негіздері, әдістері және жетістіктері

Жоспар
І. Кіріспе
ІІ. Негізгі бөлім
2. 1. Жеке өсімдік дамуының малекулалық негіздері
2. 2. Селекцияның міндеті және селекциямен алынған материалды бағалау жолдары.
2. 3. Селекцияның әдістері мен жетістіктері
ІІІ. Қорытынды
IV. Пайдаланылған әдебиеттер
Кіріспе
Өсімдіктер әлемінде көп кездесетін полиплоидтық және гетерозис құбылыстары генетикалық факторларға жатады. Өсімдік клеткаларында хромосомалар санының еселеніп көбеюі полиплоидия деп аталады. Жыныс клеткалар ядросында хромосомдар жиынтығының неше рет қайталанатын шығын көрсететін сан - плоидтылық - жеке түрдің өзіне тән белгісі болып есептеледі.
Өсімдіктердің дене (соматикалык) клеткаларында, әдетте, хромосомалар саны екі еселенген (2п) болып келеді. Әр жұптағы хромосоманың біреуі - аналық, екіншісі аталыкқа жатады. Яғни, бұл жағдайда гаплоидты (жалқы) жыныс клеткалардан (гаметалардан) бір-бірден қосылғаи хромосомдар жиынтығы қалыптасады. Жынысты жолмен көбейтетін организмдердің көпшілігі диплоидтарға жатады.
Полиплоидты организмдерде жеке хромосом үш рет (Зп) - тримлоидты, төрт рет (4п) - тетраплоидты, бес рет (5п) - пентаплоидты, т. б. клеткалар пайда болуы мүмкін.
Полиплоидты өсімдіктерлің клеткаларының, жеке мүшелерінің (жапырақ, гүл шоғырлар, жемістер), тұтас денесінің шамадан тыс ұлғайып өсуі жиі байкалады. Көп жағдайда белгілі химиялық қосындылардың мөлшері көбейіп, гүлдену және жемістену мерзімдері өзгереді.
Полиплоидтылыққа жоғары және төмен температура, иондаушы саулелену, химиялық заттар, сондай-ақ клеткалардың физиологиялық күйінің өзгеруі себепкер болуы мүмкін.
Түрліше әдістермен будандастыру арқылы алынған будандардың (гибридтердің) бірінші ұрпағының өсу қарқындылығының жоғарылап, өлшемдерінің ұлғайып, жемістілігінің көбеюі гетерозис, немесе -будан күші» деп аталады.
Әдетте гетерозис басымдылығы шыққан тектері жағынан алшақ топтарды, түрлерді өзара будандастыру арқылы алынған, бірінші ұрпақ «кілдерінде ғана байқалалы. Одан әрі қарай екінші, үшінші, т. б. ұрпақтарда оның күші біртіндеп азайып, жойылып та кетеді. Гетерочис құбылысының негіздері туралы түрліше ғылыми көзқарастар кездеседі.
Солардың ішіндегі «аса басымдылық» немесе моногенді гетерозис деген болжамға сәйкес гетерозиготалары жекеленген гендерге лайықты белгілері гомозиготаларға қарағанда басым келеді. Басқа бір болжамдар будандардя ата-аналарына карағанда әр түрлі гендердің басым аллельдері көп болатындығына және солардың арасындағы әрекеттестіктер туралы ережеге негізделген. Жинақтаушы (синтездік) сипаттағы болжамдар ішкі және гендер аралық әрекеттестіктерге негізделген.
Жалпы айтқанда, гетерозиготалық болжам және геннің екі күйде болатындывын, өзара қосылғанда организмнің өсуі мен дамуын өзгертетіндігін уағыздайды. Ал «аса басымдылық» болжамы бойынша будандардағы аталық пен аналықтың басым гендерінің бірігуі гетерозис құбылысына себепші болады деп есептейді.
Гетерозис құбылысының егін шаруашылығында маңызы өте үлкен. Гетерозисті будандардың (астық дақылдар) түсімлілігі 20-30 % артық болады. Дегенмен, ол ауылшаруашылық тәжірибесінде кең таралған жоқ. Себебі, осы уакытқа дейін геторозис құбылысын бірнеше ұрпақта тұрақтандыру мәселесі әлі шешілмей келеді.
2. 1. Жеке өсімдік дамуының малекулалық негіздері
Соңғы кездерде тіршіліктің негізгі белгілері, соның ішінде тұқым қуалаушылық пен даму молекулалық деңгейде зерттеле басталды. Молекулалық биологияның қалыптасуына байланысты өсімдіктердің жеке даму процесін дұрыс, толығырақ түсінуге және онтогенездің жан-жақты теориясын жасауға жеткілікті мүмкіндік туды.
Өсімдіктердің дәуірден-дәуірге үздіксіз дамуының (эполюция) нәтижесінде ішкі (эндогеңді) ырғақтылық деп аталатын белгілері қалыптасты. Сонымен қатар ішкі факторлар эндогенді ырғақтылықтың мерзімді өзгеріп тұратын сыртқы орта жағдайларына үйлесімділігін (сөйкестілігін) анықтайды (күннің ұзақтығы, жарық деңгейі, температура, т. б. ) .
Молекулалық генетика мен молекулалық биологияның тәжірибелік зерттеулерінің жетістіктеріне байланысты организмдердің жеке дамуының ішкі сырлары (механизмдері туралы) ғылыми болжаулар қалыптасуда. Ондай көзқарастар тұқым қуалаушылықтың гендік теориясына негізделген.
Тіршілікке ғана тон - тұқым қуалаушылық - организмдердің полипептидтік тізбегіндегі амин қышқылдарының орналасу ретін анықтайтын ақпаратты жинақтап, сақтап және ұлпаларға жеткізу қабілеттілігі.
Тұқым қуалаушылық белгілердің ұрпаққа ауысу проңестері клетка ядросындағы храмасомалармен реттеледі. Хромосомаларда ерекше түзілімдер, химиялық жағынан дезоксирибонуклеин қышқылы болып есептелетін гендер бар, ДНК, молекуласында клетка белоктарының құрылысын, яғни организмнің барлық белгілерін анықтайтын толык ақпарат (информация) сақталады. Ұрпақтарға тұқым қуалау ақпараты ДНҚ молекуласының екі еселенуі, немесе редупликациялануы арқылы жеткізіледі. Белоктың синтезделуі рибосомаларда (полисомаларда) жүзеге асалы.
Белоктың синтезделуіне рибонуклеин қышқылдары (РНК, ) - рибосомалық (рРНҚ), тасымалдаушы (тРНК. ) және ақпаратшы, немесе матрикстік (мРНҚ) қатысады. Тасымалдаушы РНҚ амин қышқылдарды рибосомаларға ауыстырып, мРНҚ болса, генетикалық ақпаратыг ДНҚ-ның нуклеотидтік шифрына, алдымен тұрақсыз қосыңдыларға
- РНҚ-дарға, соңынан амин қышқылдар шифрына «көшірмелейді». Белок молекуласындағы амин қышқылдарының бір ізділігі (реттілігі) мРНК, молекуласындағы нуклеотидтер реттілігіне ауысуы транслация деп аталады.
Рибонуклеин қышқылдарынын барлық түрі ДНК, матрицасында РНҚ-полимераза ферментінің көмегімен синтезделеді. Содан соң тРНК, мен мРНҚ цитоплазмаға тасымалданып, рибосомалардағы белок синтезіне қатысады.
Сонымен, организмдердің тұқым қуалаушылық белгілері мынандай жолмен беріледі деп есептеледі: ДНҚ РНҚ белок зат алмасу тұқым куалаушылық қасиеттер мен белгілердің қалыптасуы. Бірақ, зерттеулердің нәтижесінде белок биосинтезінің матрицасы ДНҚ-да ғана емес. РНҚ-да болуы мүмкіндігі анықталды. Мысалы. ДНҚ-сы жоқ өсімдік вирустердіде белок РНК-да синтезделеді.
Организмдер ашық, өсіңкі жүйе ретінде қоршаған ортамен үздіксіз әрекеттестікте болады. Осы байланыстықтың арқасында олар ақпараттың қосымша қырына ие болады (6. 11-сурет) .
Өсімдіктердің дамуын меристема клеткаларынын жіктелуінен елестетуге болады. Бұл ұлпалар бөлінудің және соған сәйкес зат алмасудың нәтижесінде, ішкі және сыртқы жағдайлар әсерінен ерекшеленген ұлпаларға айналып, дамудың өсу (вегативтік) сатысынан көбею сатысына ауысады. Әрбір жеке клеткада генетикалық ақпараттың толық жиынтығы болады деп есетеледі. Яғни, жеке клетка татипотентті - өздігінен дамып, тұтас организмге айналу қасиетіне ие деуге болалы. Клеткадағы биохимиялық және морфогенетикалық процестердің-көпшілігі ферменттердің қатысуымен жүзеге асады. Клеткада іске асатын химиялық реакциялардың әрқайсысына тән белгілі ферменттің болуы - биологияның негізгі заңы болып есептеледі.
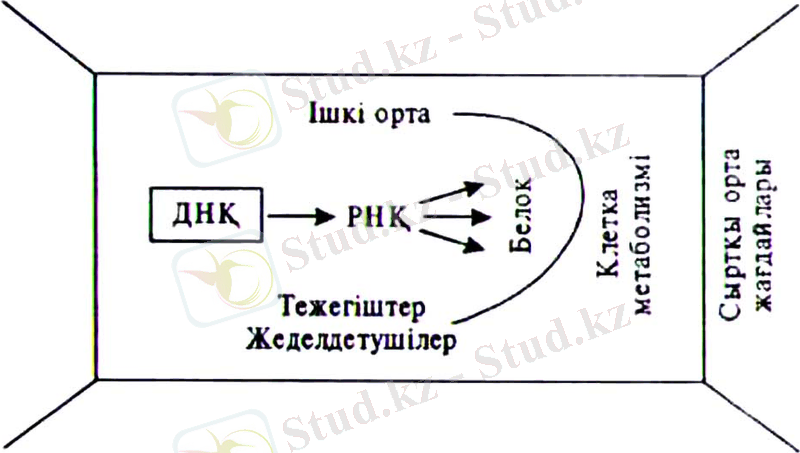
6. 11 - сурет . Клетка мегаболизміне ішкі және сыртқы жағдайлар ықпалының графикалық бейнесі (С. ИЛебедев, 1488) .
Белгілі өсімдік организміне тән зат алмасу ерекшелігі оның генетикалық тегімен анықталады. Оның ерекше ферменттгер жиынтығы да, және ферменттік процестердің ара-қатынасы да осыған байланысты. Сонымен өсімдіктердің тіршілік жағдайларының өзгеруше байланысты ішкі процестерді өздігінен реттеушілік қабілеті екі жолмен ферменттердің синтезделуін реттеу және олардың ырықтығын реттеу арқылы жүзеге асады.
Француз ғалымдары генетик Ф. Жакоб және биохимик Ж-Моно теориясына сәйкес әлде бір ферменттің синтезделуін қоздыратын, немесе тежейтін қосындылар құрылыстық гендерге екі еселенген генетикалық. өзгеру арқылы әсер етеді. Құрылыстық гендер белоктардың молекулалық құрайтын анықтайды. Полинуклеопитті тізбекте құрылыстық гендер тобымен қатар ген - жұмысшы (ген-оператор) немесе оперон деп аталатып генетикалық реттеу бірлігі де орналасады. Оперон - қызметтері жағынан өзара байланысты ферменттер тобының синтезделуін анықтайтын гендер тобы. Оперон белгілі ферменттердің синтезделуіне қажетті құрылыстық гендердің қызметін қадағалап реттейді. Ген-оператор жайындағы промотормен ДНҚ тәуелді РНК, - полимераза ферменті байланысады. Ол А және Б ферменттер синтезін кодтейтін матрица - РНҚ-ны синтездейді. Ген-оператор қызметі өз кезегінде репрессор (тежегіш) деп аталатын затқа тәуелді болады. Ол белок тектес болуы мүмкін және ген-реттеуші ықпалына байланысты пайда болады. Репрессор осы ферменттің синтезін қоздырушы, соңдай-ақ оның пайда болуын тежеуші қосындылармен әрекеттесе алады (қозу. тежеу) (6. 12-сурет) .
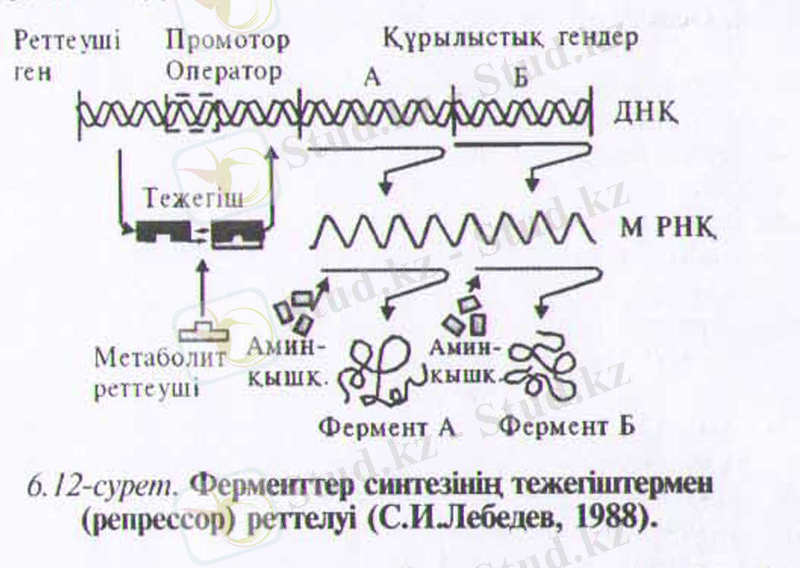
Өсімдіктер организміндегі тежегіштер (репрессор) және эффекторлар (әсерлегіштер) әсерлеріне бір-екі мысал келтірейік.
Жаңа жиналған картоп түйнектері, қажетті жағдайлар болғанымен, өнбейді. Себебі олардың көзшелері тыныштық күйде болады. Бұл кезде картоп көзшелеріндегі геном (гендер жиынтығы) толығынан тежлген күйде болатындықтан, РНҚ синтездеуге қабілетсіздіктен белок синтездеуші жүйелер де ырықсыз күйде болады. Егер көзшелер гибберелл қышқылымен немесе этиленхлоргидринмен өнделсе, онда олар өне бастайды. Бұл заттар көзше геномдарына қоздырғыш ретінде әсер етіп, тежелген күйден шығарады (дерепрессия) . Гендердін біразы гистон белоктарынан босауы мүмкін; осы ырықты гендердің ыкпалынан картоп көзшелерінің тіршілігіне кажетті перменттер синтезделеді.
Арпа дәнінің эндоспермінде а-амилаза синтезін бақылайтын ген тек тұқым өнген кезде ғана іске қосылады. Өнген ұрықта пайда болған гибберелл қышқылын а-амилазаның генін «босатып, нәтижесінде фермент синтезделеді.
Жалпы айтқанда өсімдіктердің вегетативтік өсу сатысынан көбею сатысын ауысуын, молекулалық биология тұрғысынан төмеішсгіше түиндіруте болады.
Белгілі фотопериод (жарық мерзімділік) әсерінен жапырақтарда ерекше заттар - әсерлегіштер пайда болады (стеройд тектес) . Олар тиісті клеткаларға тасымалданып, тежелген күйдегі гендерді босатып, іске қосады. Босанған гендер гүлдердін, жемістердің, тұқымдардың, т. б. қалыптасуына қатысады. Осы гендерде РНҚ синтезделіп, оның әсерінен көбею органдарының дамуына қажет ферменттер пайда болады. Мысалы, көктемде қызыл шиенің көбеюі бүршіктерінде нуклеин қышқылдарынын мөлшері, өсу бүршіктердегіге қарағанда 1, 5-2 есе көп болады. Жемістейтін бүршіктерде гистондардың мөлшері азайып, РНК, концентрациясының көбеюі тиісті белок - ферментгерлін синтезделуін күшейтеді.
2. 2. Cелекцияның міндеті және селекциямен алынған материалды бағалау жолдары
Селекцияның негізгі міндеті - аурулар мен зиянкестерге, қуатьтылық пен суыққа төзімді, дәннің сапасы жоғары, интенсивті егіншілік талаптарына сай келетін дақылдардын жана сорттарын шығарып, өндіріске енгізу.
Әр өсімдік түрінің қазіргі жағдайда ең жоғары, сапалы өнім беретіп бір тобын, ауыл шаруашылық дақылдарының сорты деп айтады. Бір сортқа жататын өсімдіктердің биологниялық шаруашылық сипаттамалары және пішіні де біркелкі болып келуге тиіс және осы көрсеткіштері бойынша бағаланады. Әр сорт белгілі бір аймаққа, сол жердің ауа райына, топырақ жағдайына толық сәйкестелген болады. Сонда ғана ол жыл сайын тұрақты өнім береді. Өнімиің сапасы да жақсы болады. Шаруашылықтарға аудандастырылатын жаңа сорт, мұнда бұрын егіліп жүрген сорттарға карағанда өнімі мол және сапасы да жоғары болу керек.
Өсімдіктердің түрлері, оның түр тармақтары бір-бірінен биологиялық сипаттарына, тиімділік қасиеттеріне, құрылысына қарай бөлінеді. Сондықтан да өсімдіктің бір түріне жататын оның тұр-тармақтарында бәріне ортақ белгілер болуы тиіс.
Жаңа сорт шығарудык екінші жолы - өсімдіктерді бірімен-бірін будандастыру. Бұл әдісті көп қолданып, жақсы сорттар шығарған селекционер ғалымдардың бірі Иваи Владимирович Мичурин.
И. В. Мичурин бақта өсірілетін жеміс-жидектердің 300-ден аса жақсы сорттарын шығарды. Ол сорттар еліміздіц көп аудандарында әлденеше мың гектар жерге егіледі. Сотрт шығаруда Мичурин өсімдікті будандастыру әдісін ғана қолданып қойған жоқ. Ол өсімдіктерді іріктеу, сұрыптау, ұластыру, оларды күту жағдайын жақсартып, өсіріп-жетілдіру әдістерін де кең қолданды.
Әріден будандастырғанда әр, түрлі өсімдіктердің туыстары арасында будандастайтын жағдайлар да байқалады. «Мұндай жағдайда В. И. Мичурин тозаң қосасын қолдануды, алдын ала өсімдіктерді вегетативті жақындастыру және аралық будандастыру сияқты ерекше, тиімді әдістерді қолдануды ұсынды.
Алғашқы әдісті қолданғанда, аталық өсімдіктің тозаңына аналық өсімдік тозанының біраз бөлігі немесе сол өсімдіктің бірнеше сортының тозаңы қосылады, мұның өзі көбінесе будандастыруды жақсартады.
Өсімдіктерді вегетативті жақындастыру әдісінің маңызы - есейген өсімдіктің (мәселен, шетен - жеміс ағаштарының бір түрі және оның жемісі) бөрікбасына бұдан көшетінің жас сатысындағы (мәселен алмұрт) қалемшесі уластырылады. Есейген өсімдіктің шырынымен қоректене отырып, сеппенің ұластырылған өсімдікпен будандасу қабілеті біліне бастайды.
Екі түрлі өсімдіктің тікелей будандастырылуынан ешнәрсе шықпаған жағдайда, Мичурин аралық будандастыруды ұсынады, мұнда будандастыруға өсімдіктін үшінші түрі енгізіледі. Өсімдіктің бірінші түрін үшінші түрімен будандастырғанда, оның «аралық» будан көшеті пайда болады, оны өсімдіктің екінші түрімен будандастырады. Осылайша жаңа сорт алынады.
ТМД-да 44 селекциялық орталық құрылды, 100 үстінде сортты тұқымдық бірлестігі, қала берсе сорттың тұқым өндірісін құру толығымен аяқталды.
Ғылыми-зерттеу институттары, селекциялық сортты тұқымының орталық бірлестігі облыстық ауыл шаруашылық тәжірибе станциялары сортты шығарғаннан кейін, өндірістік жағдайда тағы бірнеше жыл тексеріледі, сынайды.
Жаңа шығарылған сорттарды сынау, тексеру, содан кейін аудандастыру мәселесін шешу, арнаулы мекемелерге тапсырылады.
Қазіргі кездегі өсімдіктер селекциясының мақсаты -әр түрлі аймакқа арналған түрлі дақылдарды мүмкікдігінше көбірек бейімдей түсу, яғни белгілі бір генотип пек сортқа өсетін ортаның абиотикалық және биотикалық факторлары әсерінің үйлесімділігін барынша арттыра түсу болып табылады.
Селекцияның алғашқы қадамы Кіші Азия жерінде басталғаны белгілі. Сол алғашқы қадамның нәтижесінде бұрын аң аулап, жабайы өсімдіктер теріп, тіршілік еткен халық біртіндеп отырықшылыққа көшеді. Венгрияның белгілі селекционер ғалымы Лелли Кіші Азиядағы халықтар мәденнетінің дамуына, олардың санының тез өсуіне бұл жерде астық тұқымдас өсімдіктерлің екі туысы - бидай мен арпаны өсіру негізгі себем болғанын зерттеп анықтаған. Бұл жердің халқы осы екі дақылды ең алғаш қолдан өсіруді үйренеді, сөйтіп диқандыққа қолдары жетеді. Осының нәтижесінде тұрақты мекендер орнап қалалар салынады.
ТМД-де ауыл шаруашылығы саласындағы ғылыми-зерттеу мекемелері, селекциялық орталықтар соңғы жылдары мол түсімді және дан сапасы жоғары интенсивті бірнеше ауыл шаруашылығы дақылдарының сорттарын өсіріп шығарды.
Астық түсімі сол дақылдык сортына байланысты болатыны жөнінде еліміздің көрнекті ғалымы академик Н. И. Вавилов Совет өкіметі орнаған алғашқы жылдары-ақ айтқан болатын. Соған орай біздің ғалымдарымыз қазіргі кезде сорттарды аудандастыру нәтижесінде дәнді дақылдардың гектар өнімділігін 25 центігерден 40 центнерге дейін арттыруға болатынын дәлелдеп берді.
Қазақстан жерінде күні бүгінге дейін ауыл шаруашылығы дақылдарының 9300 сорты, оның ішінде республика селекционерлері шығарған 830 сорты апробациядан өткізілді. Тек соңғы 12 жыл ішінде 37 дақылдың 127 сорты аудандастырылды.
Әрине жыл өткен сайын жақсы сорттардыц орніііна одаң да тәуір сорттар шығарылып жатады. Жақсы сорттар өндіріске енгізілген санын гектар берекесі артып, жаллы өнім де көбейе береді. Сондықтан да совет ғалымдары дәнді дақылдардың, оның ішінде бидайдын жаңа сорттарын шығаруға байланысты ізденуді тынбай жалғастыруда.
Соңғы жылдары Ленинградтағы агрофизика ииститутындағы профессор Борис Мошков басқарған лаборатория ғалымдары бидай өсімдігінін тағы бір сырын ашты. Бірнеше жылға созылған тынымсыз еңбек пен іздеуудің нәтижесі күптеген жаңалыққа алып келді. Бидайды және басқа да масақты дәнді дақылдарды интенсивті фотосинтездеу арқылы тәжірибе жүргізгенде, олар жәқсы өсіп, өте жақсы вегетациялық масса беретінін, ал дән саны аз болатынын көрсетті. Ал орташа фотосиитезделген кезде керісіншс, бір түп бидайдың 200 г массасы алынса, әр масақ 150-180 дән берді. Мұндай эксперименттің нәтижесі болашақта себу нормасы аз болатын жаңа бидай сортын шығару жұмысын бастауға негіз болмақ.
Еліміздің шығысындағы негізгі астықты аймақ болып есептелетін Қазақстанның ауыл шауашылығы ғылыми-зерттеу
2. 3. Селекцияның әдістері мен жетістіктері
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz