Наубайханалық ашытқы өндірісінің биотехнологиясы: технологиялық схема, сапа көрсеткіштері және қауіпсіздік талаптары

«Нан ашытқысы өндірісінің биотехнологиясы»
дипломдық жұмысына
АННОТАЦИЯ
Дипломдық жұмыста нан ашытқысы өндірісінің биотехнологиясы жасалған. Наубайханалық ашытқы өндірісінің негіздері талданды. Ашытқы жасушасының құрылысы мен химиялық құрамы келтірілді. Наубайханалақ ашытқы өндірісінің технологиясын құрастырылды. Наубайханалақ ашытқының және мелассаның сапа көрсеткіштері анықталды. Қажетті технологиялық есептеулер жасалды. Еңбекті қорғау және тіршілік қауіпсіздігі мәселелері қарастырылды.
АННОТАЦИЯ
дипломной работы на тему «Биотехнология производства хлебопекарных дрожжей»
Дипломная работа посвящена разработке биотехнологии хлебопекарных дрожжей. Сделан анализ основ производства хлебопекарных дрожжей. Даны строение и химический состав дрожжевой клетки. Разработана технологическая схема производства хлебопекарных дрожжей. . Даны показатели качества хлебопекарных дрожжей и мелассы. Рассмотрены вопросы охраны труда и безопасности жизнедеятельности.
Summary
the thesis on the subject "biotechnology of production of baker's yeast"
In the thesis it is devoted to development of biotechnology of baker's yeast. the analysis of bases of production of baker's yeast is made. the structure and a chemical composition of a barmy cage are given. the technological scheme of production of baker's yeast is developed. indicators of quality of baker's yeast and molasses are given. questions of labor protection and health and safety are considered.
мазмұны
Жұмыс уақыты мен қондырғылардың
санын есептеу
Кіріспе
Ашытқы дегеніміз бүршіктенетін немесе бөлінетін бір жасушалы саңырауқұлақ, өлшемдері бойынша микро ағза болып табылады. Микроағзалар алғаш рет ХVII ғ. ортрасында Левенгук ашқан еді. Қазіргі түсініктер бойынша ашытқылар жеткілікті дәрежеде бір жасушалы ағза болып табылады. [1]
Ашытқы өндірісі кептірілген және пірестелген ашытқылар өндіреді. Негізінен ашытқыларды наубайхана, кондитер, консерві өндірістерінде қолданылады. Сонымен қатар, ашытқыларды дәрумен өндіру өндірісінде Д және В 2 алу үшін, медицина саласында емдік препараттар, нуклейн қышқылдарын, түрлі ферменттерді алу үшін микробиология саласында қоректік орта дайындау үшін, ауыл шаруашылығында жас малдарды өсіруге, құс фермалары мен балық шаруашылығында кеңінен қолданылады. Ашытқы өндіру зауыттары өткен ғасырдың ортасында пайда болды. Ашытқы өндіру үшін негізгі шикізат ашытқы жасушаларының өсіп дамуына қажетті заттар мен бай астық дәні бидай қолданылды. ХХ ғасырдың басында наубайханалық ашытқыларды өндіруге арналған бидай құны жоғары болғандықтан, қант қызылша өндірісінің қалдығы мелассамен алмастырылды. Мелассамен қатар спирт алынады. ХХ ғасырдың ортасында ашытқы өндірісінің технологиясы біршама өзгеріске ұшырады. Бұл өзгерістерге ғалымдардың наубайханалық ашытқыларды мелассалық ортада өсіру технологиялық режимдерінің теориялық негіздеуі және қондырғылармен жабдықталып, прогрессивті технологиялар мен қамтамасыз етілді. Ашытқыларды өндірудің үздіксіз технологиялық нобайлары өндірістік өнімділікті 15-20% ға өсіруге мұмкіндік берді. Қазіргі таңда өндіріске концентрленген ортада ашытқыларды алу перспективті режимдері өндірілуде.
Концентрленген орталарды қолдану нәтижесінде ашытқы өсіру аппараттардың биомасса түсіру салыстырмалы көлемі 5кг(м3х сағ) тың орнына 10-17 кг (м3 х сағ) құрады. Алынған дайын өнім сапасы да біршама жақсара түскен. Өндіріске енгізілген ашытқыларды өндіру жаңа әдістері өндірістің өнімділігін жақсартып ғана қоймай, сонымен қатар шығарылған дайын өнім престелген және кептірілген ашытқылардың сапасын жақсартып, шығымын жоғарлатады су ресурстарын үнемдеуге мүмкіндік береді. Ашытқы өсіру аппараттарын, ауа шашырату жүйелерін, кептіру агрегаттарын жетілдіруге және ашытқы өндірісін автоматтандыру үнемі қолға алынып отырады. Жоғары өнімділіктегі жаңа ашытқы рассалары селекцияланып, арнайы рассалар кептірілген ашытқылар үшін дайындалады. Ағымды суларды ластамауға арналған ашытқыларды өсіру режимдері құрастырылуда. Ашытқы өндірісінің қызметкерлері бражкада дайындалатын ақуызды концентрат ойлап тапқан. Ашытқы өндірісінің техникалық-экономикалық көрсеткіштерін жоғарылату мақсатында қазіргі заманғы қондырғыларды, кептірілген және престелген өнімдерді өндіру технологиялық режимдерін, шикізаттарды есептеу, нормадан ауытқыған кезде технологиялық процесті түзете алуын әр технолог білуі шарт.
Дипломдық жұмыстың мақсаты - наубайханалақ ашытқы өндірісінің технологиясын құрастыру болып табылады.
Осы мақсатты іске асыру үшін алға келесі міндеттер қойылды:
- наубайханалық ашытқы өндірісінің негіздерімен танысу;
- ашытқы жасушасының құрылысымен, химиялық құрамымен танысу;
- наубайханалақ ашытқы өндірісінің технологиясын құрастыру;
- наубайханалақ ашытқының және мелассаның сапа көрсеткіштерімен танысу;
- қажетті технологиялық есептеулер жасау;
- еңбекті қорғау және тіршілік қауіпсіздігі қарастыру.
1 НАУБАЙХАНАЛЫҚ АШЫТҚЫ ӨНДІРІСІНІҢ НЕГІЗДЕРІ
1. 1 Өндірістің микробиологиялық негіздері
Микроскопиялық саңырауқұлақтар. Амилаза алу үшін Aspergileus тектес микроскопиялық саңырауқұлақты қолданды. Аспергильдеу үшін қоректендіру ортасына көміртегі, азотты және минералды заттар енгізіледі.
Амилазаны жинақтау үшін онда декстрина немесе мальтоза болуы тиіс, азот көздеріне ақуыздар олардың гидролизаты, аммоний тұздары мен нитраттар жатады.
Ашытқылар. Қантты ашыту үшін спирт өнеркәсібінде Saccharamyces cerevisiol түріндегі ашытқыларды қолданады. Осындай типтегі бір - бірінен кейбір белгілері бойынша ерекшеленетін бірнеше түрлі жасушалар пайдаланылады.
Дән, картоп шикізатын қайта өңдейтін спирт зауыттарында мальтоза, сахароза, фруктоза жақсы ашытатын, бірақ соңғысы декстринді ашытпайтын ашытқыларды қолданады.
Ашытқы саңырауқұлақтары - бір клеткалы қозғалмайтын және бактериялардан шамамен алғанда он еседей ірі микроорганизмдер. Табиғатта кең тараған. Жасуша пішіні әртүрлі: дөңгелек, сопақша және таяқша тәрізді болады. Ашытқы саңырауқұлақтары жасушасының мөлшері 8-10 микронға тең [2] .
Оларда қозғалу органеллалары болмайды. Жасуша сыртында қабығы бар. Цитоплазмада ядро, ваколя және басқа да (май, гликоген, валютин) заттары кездеседі. Ашытқы саңырауқұлағын адам қолдан өсіріп, өз шаруашылығында пайдаланады. Ал, табиғатта жабайы ашытқы саңырауқұлақтары да болады. Олар ауылшаруашылық өнімдерін зақымдап, едәуір зиянын тигізеді. Ашытқы саңырауқұлағының адам баласына пайда келтіретін түрлерін біз мәдени ашытқы саңырауқұлақтары деп атаймыз. Ашытқы саңырауқұлақтары өнеркәсіпте кең қолданылады. Олар қантты ашытып, көмірқышқыл газы мен спирт түзеді. Олардың бұл қасиеті нан ашытқы өндірісінде және спирт өндіруде, түрлі шараптарды, сыраларды, сүт тағамдарын даярлауда қолданылады. Ашытқы саңырауқұлақтарында ақуыз және витаминдер (В. Д. Е) көп болады, сондықтан оларды қазір тамақ және мал азығын алу мақсатында кеңінен қолданады.
Ашытқы саңырауқұлағы көбінесе бүршіктену арқылы көбейеді. Бұлар спора түзу және жәй бөліну арқылы сирек көбейеді. Олардың кейбір түрлері жыныстық жолмен көбейеді.
Бүршіктеніп көбейгенде, алдымен аналық жасушадан төмпешік пайда болады да, кейіннен ол үлкейіп бүршікке айналады. Одан кейін жас жасуша аналық организмнен мүлдем бөлініп кетеді. Қолайлы жағдайда бүршіктену екі сағатқа созылады. Көмірсу мен азотты қоректік затқа бай ортада ашытқы саңырауқұлағының бүршіктенуін жәй биологиялық микроскоптармен де көруге болады.
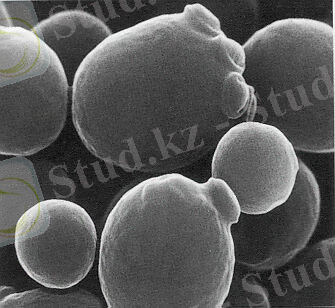
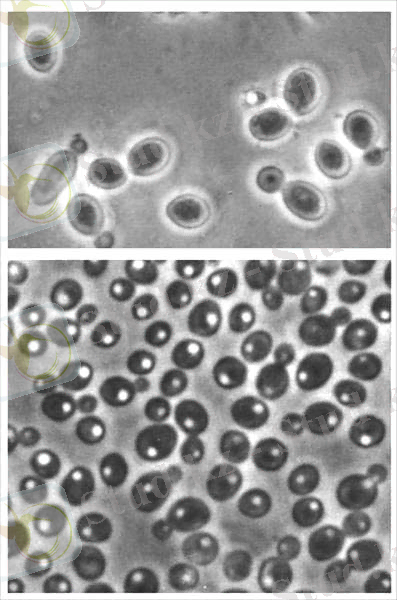
Сурет 1. Ашытқы саңырауқұлақтары
Спора арқылы көбею оларда жыныстық және жыныссыз жолдармен жүреді. Ашытқы саңырауқұлақтары жасушасындағы споралардың саны екіден он екіге дейін барады. Жыныссыз жолмен спора пайда болғанда вегетативтік жасушалар ұсақ бөлшектерге бөлінеді де олардың әр қайсысының сыртында қабық пайда болады. Ал жыныстық жолмен спора пайда болған екі жасуша қосылады да оның сыртында қабық пайда болады. Споралардың пішіні дөңгелек немесе сопақша болып келеді.
Ашытқы саңырауқұлақтардың систематикасы көбею тәсілдері мен физиологиялық қасиеттеріне негізделген. Олар екі тұқымдасқа бөлінеді: сахаромицеттер және сахаромицет еместер.
Сахаромицеттер. Бұларға мәдени ашытқы саңырауқұлақтары жатады. Олар бүршіктену және спора түзу арқылы көбейеді. Сондықтан бұларды нағыз ашытқы саңырауқұлақтары деп атайды. Мәдени ашытқы саңырауқұлағынан нан, шарап, сыра, ашытқы саңырауқұлақтары жатады.
Өндірісте, әсіресе олардың сахаромицес церевидзе және сахаромицес элипсойдеус деген түрлерінің маңызы зор.
Сахаромицес церевидзе жасушасы шар немесе жұмыртқа тәрізді. Олар шарап спиртін алу үшін, сыра қайнатуда және нан ашытуда кеңінен қолданылады. Бұлардың бегілі бір температурада және жағдайларда тіршілік ететін жеке топтары - рассалары бар. Ал сахаромицес эллипсойдеустің жасушасы эллипс тәрізді, шарап өнеркәсібінде қолданылады, олардың кейбір расалары шараптағы хош иісті түзуге тікелей қатысады.
Сахоромицет емес ашытқы саңырауқұлағы негізінен жалған ашытқы саңырауқұлақтар. Олай деп аталатын себебі: ашытқы саңырауқұлақтарының спора түзуге қабілеті болмайды, бүршіктену арқылы ғана көбейе алады. Бұлардың көпшілігі әртүрлі өндірістегі өнімдерді зақымдайды. Дегенмен олардың ішінде шаруашылық үшін маңызды туыстары бар. Олар: торула және микодерма. Торула туысына жататын ашытқы саңырауқұлақтары шар тәрізді келеді және ашыту процесі барысында азғана мөлшерде спирт түзеді. Торула айран деп аталатын өкілі қымыз және айран сияқты сүт тағамдарын даярлауда пайдаланылады, ал торула утилис-тағамдық және мал азығы ашытқы саңырауқұлақтардың жасушасы ұзынша. Олар спирт түзе алмайды. Бірақ ортадағы бар спиртті және органикалық қышқылдарды су мен көмірқышқыл газына дейін тотықтыра алады.
Құрамында спирті бар ішімдіктердің бетіне қонса, микодерма қатпарланған қабықша түзеді де, оның иісі мен дәмін бұза бастайды. Сонымен қатар микодерма сүт тағамдарын, тұздалған жидектерді бүлдіріп, сірке және нан ашытқы саңырауқұлағын жасайтын өндірістерге өте зиянын тигізеді.
1. 2 Ашытқы жасушасының құрылысы
Ашытқылардың жіктелуі [3] . Ашытқылардың жіктелу негізінен олардың спора түзу қабілеттілігі жатады. Спора түзетін немесе түзбейтін, ашытқыға ұқсас саңырауқұлақтар. Спора түзетін және ашытқыға ұқсас саңырауқұлақтардың көбеуі. Sacchavomycetes классындағы бүршіктену арқылы көбейетін спора түзетін ашытқылар. Спора түзетін саңырауқұлақтар тұқымдастары: Sacchavomycetaclae, Sacchavomycodaceae, Schizosacchavomy cetaceae. Gziptococcaceae тұқымдас бүршіктенбейтін, ашытқылар және ашытқы тектес саңырауқұлақтар. Жемдік ашытқылар өндірісінде аспорогенді саңырауқұлақтарды қолдану. Наубайханалық ашытқылардың мәдениеттері мелассалық орталарға тұрақтылығы гинетакалық белсенділігін анықтайды., өсіру салыстырмалы жылдамдығына ие болу тиіс және құрамындағы ферменттер ß - фруктофура нозидаза және λ - глюкозидаза белсенді жоғары болады. Наубайханалық ашытқылардың жасушалары ірі, шаң-тозаңды болады. Наубайханалық ашытқыларды өндіру үшін келесі расалар мен штаммдар қолданылады: Толеская 7, ЛБД - 1, Одессалық 14, л - 441, А-1, киевтік 21, гибридті расалар. Ашытқы жасушаларының құрылымы. Ашытқы жасушасы қабықтан, цитоплазматикалық мембрамадан және цитоплазмадан тұрады.
Морфологиясы. Ашытқы жасушасы жасушалы қабырғадан, цитоплазмалық мембранадан, протоплазмадан, цитоплазмалық қосылыстар- митохондриядан, рибосомадан, ядродан, вакуоля және қоректік заттар волютин, гликоген, майлы тамшылардан тұрады.
Жасушалы қабырға жасушаның негізгі құрылымдық элементі болып табылады. Қабырға қалыңдығы 1500-2500нм құрайды. Оның құрамына 90% көмірсулар 10% ақуыздар, липидтер, хитиннен тұрады. Орындайтын функциясы жасуша жасуша формасын бір қалыпты ұстап тұрады.
Цитоплазмалық мембрана жасушалы қабырғадан кейін орналасады. Қалыңдығы 75-80нм, ақуыздармен липидтерден тұрады. Орындайтын функциясы жасушаның зат алмасуын реттейді.
Цитоплазма (протоплазма) жартылай сұйық фаза, негізінен коллойдтардан тұрады. Онда түрлі жасушалы құрылымдар орналасады-митохондриялар, ядро, қоректік заттар грануллалары, вакуолялар болады.
Рибосомалар -бұл ұсақ гранулалар олардың құрамына 58%ақуыз 42% рибонуклейн қышқылы (РНК) ақуызбен байланысқан түрінде болады. Бұл гранулаларда ақуыз синтезі жүреді. Бұларды ақуыз фабрикасы деп атайды.
Митохондрия - бұл бөліктер гранула немесе шыбықты жіпше түрінде болады. Олар ақуыздардан, липидтерден фосфолипидтерден, майлы қышқылдардан, ферменттер мен ДНК дан тұрады. Бұл бөліктер де энергиямен жасушаларды қамтамасыз ету процестер жүреді. Бұл бөліктерді жасушалардың энергетикалық цехтары деп атайды.
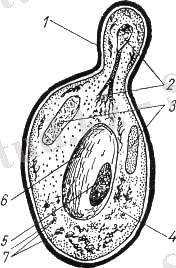
1 - қабықша; 2 - бөлінетін ядро; 3 - гликоген; 4 - цитоплазма; 5 - волютин; 6 - вакуоль; 7 - митохондрии
Сурет 2. Бүршіктелген ашытқы клеткасының құрылымы
1. 3 Ашытқылардың химиялық құрамы
Наубайханалық ашытқылардың химиялық құрамы мәдениеттеу жағдайына, қоректік ортаның құрамына жасушалардың физиологиялық жағдайына байланысты әр-түрлі болады. Престелген ашытқыларда 70-75% ылғал және 25-30% қоректік заттар болады. Судың жартысы жасуша кеңістігінде болады, оны жасуша сыртындағы су деп атайды, қалған су ашытқы цитоплазмасында болады, оны жасуша ішіндегі су деп атайды. Ішкі және сыртқы ылғал ашытқыларда ашытқы өсіруге қолданылатын рассаларға, технологиялық режимге және өсіру мен бөлу әдістеріне байланысты өзгеріп отырады. Концентрленген ортада ашытқыларды өсіргенде немесе осматикалақ белсенді заттар (мыс: натрий хлориді) қосқанда ашытқылардағы жалпы ылғал ішкі жасушалы ылғалдық азаюына байланысты төмендейді.
Ашытқылардың құрғақ заттары көміртегіден, сутегіден, оттегіден, азоттан және басқа да элементтерден тұрады. Ашытқылардың құрғақ заттардағы элементінің құрамы (%) ашытқылардың құрғақ заттары:ақуыздар және құрамында азот болатын заттар 50% майлар1, 6%, көмірсулар 33, 2%, клечатка 7, 6%, күл 7, 6% . Бірақ бұл құрам тұрақты емес және одан көп болуы мүмкін.
Ашытқылардың құрамында азотболатын заттар негізінен ақуызбен(63, 8%) нуклеопротеидтермен(26, 1%) алеидтер және пептондар (10, 1%) көрсетілген, сонымен қатар құрамында: көміртегі 53%; сутегі 7, 1%; азот 15, 7, оттегі 22, 5% және фосфор 1, 7% болады.
Ашытқылардың аминқышқылды құрамы А. С. Коновалова ұсынғаны бойынша(%) келесі түрде болады.
Аминқышқылды құрамы ашытқылардың физиологиялық жағдайына тікелей байланысты. Құрғақ заттарға қарағанда азотты емес заттар 37% құрамды, оның ішінде 32-33% көмірсулар, 4-5% май. Көмірсулар полисахаридтер түрінде болады:гликоген, галоза, глюкан, хитин.
Гликоген ең алғашқы рет 1885 жылы табылды. Ол жасушаның қосымша қоректік көмірсуы болып табылады. Құрамы және құрылымы бойынша гликоген жануар тектес гликогенге ұқсас. Гликоген ашытқы жасушаларында цитоплазмада орналасады және кодпен болғанда гранула түрінде анықталады. Ашытқылардан гликогенді алу қиын, себебі тығыз глюкан мембраналарымен қорғалған. Бұдан басқа ашытқыларда гликогеннің бірнеше фракциясы кездеседі:сілтіде еритін, сірке қышқылда еритін және хлорлы қышқылда еритін. Сілтіде еритін фракция цитоплазма компоненті, ал қышқылда еритін фракциялар жасушалы қабырға маннан мен байланысқаны белгілі.
Трегалаза ашытқы жасушасының екінші қосымша қоректік көмірсуы. Бұл екі глюкопиранозды қалдықтардан тұратын редуцирленбейтін дисахарид болып табылады. Ашытқылардағы трегалаза алғаш рет 1825 жылы Кох ғалымы тапқан. Ашытқы жасушасында трегалоза бос дисахарид күйінде болады деген пікір айтылған.
Маннан жасушаның түріне полисахарид түрінде болады. Ол ашытқылардың жасушалы қабырғасында болады және негізінен фосфат, глюкоза, глюкозамин, аминді азот аз мөлшердегі қоспасымен маннозадан тұрады.
Глюкан -полиглюкозид болып табылады. Оның құрылымы әлі анықталмаған. Бірі глюкан молекуласы ажыратылмайтын құрылымда десе, енді бірі қарсы пікірде. Электронды микроскопия және рентгенографиялық флорреакция көмегімен глюкан фибриллярлы құрылымға ие және ашытқы жасушасының формасына, жасушалы қабырғаның негізгі компоненті екені анықталған.
Хитин-N ацетилглюзамин қалдықтарынан тұратын полисахарид. Бұл химиялық инерттілігі жоғары болуымен ерекшелінеді және қиын анықталады. Ашытқы құрамындағы мөлшері аз, негізінен жасушалы қабырғада болуы мүмкін.
Л. Эблом және Е Сталпе ашытқыларды сақтау барысында көмірсулардың өзгеру динамикасына бақылау түрінде зерттеу жүргізді. Анықталғандай сірке қышқылында еритін трегалоза және глигоген мөлшері төмендеп, ал гликоген мен глюканның басқа фракциялары өзгеріссіз қалғаны байқалады. Бірақ маннан алғашқы күнде жоғарылап соңынан төмендеген. Жүргізілген зерттеулер негізінде ғалымдар келесі қорытындыға келген, жасушалардың энергетикалық қорегі сірке қышқылында еритін трегалаза және гликоген болып табылады, ал қалған көмірсулар энергия көзі ретінде қолданылмайды және жасушаның құрылымдық элементтері болып табылады.
Көптеген зерттеушілердің пікірінше, құрамында қоректік көмірсулар мол болатын ашытқылар ұзақ уақыт сақталады. Х. Суомалайнен атақты ғалым зерттеуі бойынша ауа-ағынды әдіс бойынша өсірілген ашытқыларда көмірсулардың жалпы мөлшері гликоген 5-12%; трегалоза1. 1-8. 5% маннан 17% дейін. Сақтау барысында қосымша көмірсулар мөлшері ашытқыларда төмендегі ғалымдармен дәлелденген. Н. Н Бочарова және В. Г Черный.
Мұндай ауытқулар ашытқыларды өсіру жағдайына байланысты болуы мүмкін. Осы ғалымдар ашытқыларды ауа-ылғалды әдіспен өсіру прцесінде көмірсулар метоболизімінде зерттеледі. Процес басталған да трегалаза мөлшері төмендеп, ашытқылардың интенсивті көбею кезінде минималды болатыны және процес аяқталғанда трегалаза мөлшері жоғарылайтыны анықталған. Ашытқыларды трегалаза мөлшері олардың тұрақтылығымен байланысты:трегалаза көпболған сайын, ашытқылардың тұрақтылығы арта түседі.
Ашытқылардың майлы заттары нейтралды майлардан, майлы қышқылдардан, май тәріздес заттар: липоидтер және стеролдардан тұрады. Престелген ашытқыларда 2% дейін май болады. Май негізінен қаныққан қышқылдардың бірінші қатары:пальметин 75% және стеарин 25% тұрады. Ғалымдар басқа да қышқылдарды тапқан:лаурин және олейн. Бұдан басқа ашытқы құрамында липоидтер, лицитинге жататын глицериннен тұратын фосфатидтер, фосфор қышқылы, холин және майлы қатардағы түрлі қышқылдар болады. Ашытқы майының құрамына жуылмайтын эргостерин кіреді, оның мөлшері шамамен ашытқылардың түріне және мәдениеттеу жағдайына байланысты, жалпы май мөлшерімен есептеген де 2% құрамда. Соңғы жылдары эргостеринге ашытқылардың құнарлы заттарды бағаламауы көп көңіл бөлінуде себебі; D провитамин болып табылады, ультра күлгін сәулелермен сәулелендіру кезінде ашытқылардан осы дәруменге ауысады.
Ашытқылардың майлы заттары жасуша протоплазмасының негізгі бөлігі болып табылады. Майлы заттардың молекулалары протеиннен мицелалары арасында жайылатын, таяқша тәрізді формадағы майлы мицелалармен байланысады. Кей жағдайда олар соңғы күрделі қосылыстар(мепопратиндер) түзіп, жасушаның негізгі құрылымдық материалы болып табылады.
Протоплазмада байланысқан майлардан басқа кей ашытқы соңырауқұлақтарда тамшы түрінде бос майлы заттар болады, олар арнайы бояулармен жақсы боялады.
Ашытқылардың күлі құрғақ заттарға есептеген де шамамен 6, 5-12, 0% құрайды. Кұл құрамы (%) көрсетілген. Мәдениеттеу жағдайына байланысты өзгеріп отырады.
Ашытқылардың күлі жартысы фосфордан тұрады. Ашытқылардағы фосфор қышқылының үлкен бөлігін органикалық қосылыстармен байланысқан сахаромициттердегі P 2 О 5 - мөлшері3, 8 ден 4, 4% аралығында болады. Күл құрамында магнии, натрии, кальцииге қараған да каллий мөлшері көп болады. Наубайханалық және сыра сахаромициттерінде күкірт мөлшері 0, 17-0, 20 % құрайды. Минералды заттар аралық мицелярлы суда еріп, жасуша цитоплазмасының биодинамикасында үлкен рөл атқарады. Натрий, калий, кальций, магний, теміркатиондары және хлор, фосфор амиондары маңызды болып есептеледі.
Ашытқыларда дәрумендердің бір қатары болады-В 1 , В 2 ; В 3 ; В 5 ; В 6 ; D дәрумендері жасушаның жасушаның биохимиялық процестерінде маңызды рөль атқарады. В кешендегі көптеген дәрумендер ашытқы жасушасының ферментті жүйесінің құрамына кіреді, жоғарыда келтірілген дәрумендерден басқа наубайханалық ашытқылар құрамына 1 гр құрғақ заттарға 8-95 мкг мөлшерінде парааминобензойлы қышқылы және 1гр құрғақ затқа 19-35 мкг фоли қышқылы болады.
Парааминобензойлы қышқыл фоли қышқылы бөлігі және өзі витамин ретінде әсер етеді. Бұл қышқылдар нуклейн қышқылдарының синтезін катализдейтін ферменттер құрамына кіреді.
Ашытқылар тіршілігінде биотин маңызды болып есептеледі. Сахаромициттер сыртқы ортадан биотинді синтездейалады. Сондықтан ашытқылар мәдениеттелетін қоректік ортаның құрамына өсудің маңызды факторы ретінде кіруі тиіс. Бұл дәруменнің мөлшері 1 гр құрғақ заттарға 1, 8 мкг. Биотин дәрумені карбоксилаза ферментінің құрамдас бөлігі екені анықталған, оның белсенді тобында болып, пирожүзімді қышқыл карбоксимирлену кезінде щавель- сірке қышқылына айнала отырып, азотты алмасуда және СО 2 нің сіңірілуінде қатысады.
Биотин -тұрақты зат. Термиялық өңдегенде, оттегімен және араластырылған қышқылдармен сілтілер мен әсер еткенде биологиялық белсенділігі төмендейді. Биотин тек концентрленген сілтілер мен сутегі перекись ерітіндісімен өңдегенде ғана ыдырайды.
Ашытқылардың кейбір түрлерінде өсудің басқа деңгейлендіргіші- мезоинозит болады. Бұл алты атомды циклды спирт инозитінің изомері болып табылады және 1 г құрғақ заттарға 270мкг көлемінде болады.
Анықталғандай ашытқылардың өсуіне инозит, пантотен қышқылы, аниврин, пиридоксин, параааминобензой қышқылдары әсер ететіні белгілі.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz