Ген экспрессиясы және эукариоттық транскрипцияның механизмдері мен реттелуі

Қазақстан - Ресей
Медициналық Университеті

Тексерген:
Орындаған:
Тобы:
Алматы - 2018ж.
Жоспар:
І. Кіріспе.
ІІ. Негізгі бөлім:
1. Геннің экспрессиясы
2. Транскрипция
А) Прокариоттардағы транскрипция
Б) Эукариоттардағы транскрипция
3. Трансляция
А) Рибосомалар
Б) Генетикалық код
В) Тасымалдаушы РНҚ-лар(т-РНҚ)
Г) Белоктың биосинтезі
ІІІ. Қорытынды.
ІV. Пайдаланылған әдебиеттер.
Кіріспе.
Ағзаның тұқым қуалайтын факторларын 1909 жылы В. Т. Иогансен «ген» деп атады.
Т. Морганның «Ген туралы теория» атты шығармасында (1926ж. ) берген анықтамасы бойынша:
Ген - хромосоманың бөлігі - бір белгінің жарыққа шығуына жауап беретін, бөлшектенбейтін, тұқым қуалау қызметінің, мутацияның және рекомбинацияның бірлігі деген.
Гендер хромосомада бір сызық бойында орналасқан және өз қызметін бір - біріне тәуелсіз атқарады.
Қазіргі ген теориясы - молекулалық генетиканың туындысы. Ал геннің құрылымы мен қызметін анықтауға мүмкіндік берген А. С. Серебровский, Дж. Бидл және Е. Татум, С. Бензердің жұмыстары.
Сол жұмыстардың нәтижесінде ген теориясының қағидасы қалыптасты, онда:
Ген деп, бір полипептидтік тізбектің синтезіне жауап беретін ДНҚ молекуласының бір бөлігін айтады.
Генетикалық материалдардың берілуі келесі процестер арқылы жүреді:
репликация, транскрипция, трансляция
ДНҚ ДНҚ РНҚ Белок
Сонымен қатар, тіршілікке қажетті заттар түзу үшін, жасушада генетикалық материалдардың жүзеге асырылуы ( экспрессия ) жүреді.
Ал геннің транскрипциясы мен трансляциясын біріктіріп, геннің экспрессиясы деп аталады.
Геннің құрылысын зерттеу молекулалық биология мен гендік инженерияның жетістіктеріне негізделген.
Осы зерттеулер нәтижелерінің бірі -прокариоттар мен эукариоттар гендерінің құрылысындағы айырмашылықтың ашылуы.
Прокариоттар гендері тек қана ақпараты бар (экзондар) бөліктерден тұрады.
Эукариоттар гендері бірінен соң бірі орналасқан ақпараты бар (экзондар) және ақпаратсыз (интрондар) бөліктерден тұрады.
Эукариоттар гендері экспрессиясының реттелу түрлері:
1 Ағзаның қоршаған орта өзгерістеріне бейімделуін қолдайтын уақытша реттелу;
2 Жасушалардың дифференциялануын және ұлпалардағы ақуыздар құрамының әртүрлілігін анықтайтын тұрақты, ұзаққа созылатын реттелу.
Реттелудің екі түрі де генетикалық ақпараттың іске асырылуы кезіндегі түрлі деңгейлер байқалайды: гендік, транскрипциялық, посттранскрип-циялық, трансляциялық және посттрансляциялық.
Эукариоттар организмінде гендердің реттелуі күрделі процесс. Өйткені, эукариоттарда ядро қабығының болуы, хромосома санының көп болуы, гендердің өзара әрекеттесуі, құрылымдық гендердің транскрипциясында индуктор ретінде гормондардың және жүйке жүйесінің қатысуы геннің жұмысының реттелуін күрделендіреді.
Прокариот генінің экспрессиялы реттелуі Оперон моделі (1961 Жакоб және Моно)
Прокариоттардағы гендер экспрессиясының реттелуі - белоктық құрылымы және информация құрылымдық гендерде жазылған. Бір хромосоманың бойында қатар орналасқан гендер тіркес
гендер, бұл гендер активтілігі басқа бір реттеуші генніңбақылауында болады.
Прокариоттарда реттелудің екі типі бар: негативті және позитивті.
Лак-оперон - позитивті реттелу. Мысалы, ақуыз-репрессор әрдайым жұмыс халінде болады;
Триптофанды-оперон - негативті реттелу. Мысалы, репрессор белсенсіз халде болады.
ІІ. 1. Геннің экспрессиясы
Гендер экспрессиясы - генетикалық ақпараттың полипептидтер мен ақуыздарға ДНҚ-нан РНҚ арқылы тасымалдануы.
Гендер экспрессиясының дифференциалды реттелуін жүзеге асыру қабілеті прокариоттар мен эукариоттардың негізгі қасиеті болып есептеледі. Қандай гендердің экспрессиялануы керек, қайсыларына керегі жоқ екендігіне бақылау жасай отырып, сол сияқты түрлі гендер экспрессиясының деңгейін реттей келе клеткалар өз фенотипін белгілі бір сыртқы және ішкі орта жағдайларына бейімдей алады. Гендер көбінесе біртіндеп экспрессияланады. Бір геннің активациясы басқа бірнеше гендердің экспрессиясын тудыра алады. Қайсыбір гендер немесе туыстас гендердің тобы үйлесімді түрде экспрессияланады, яғни реттеуші сигналға бір мезгілде және бірдей деңгейде жауап береді. Синтездің бір белоктан басқаларына ауысуы кез - келген дамудың, мейлі ол залалданған клеткадағы вирустардың репродукциясы, бактериядағы спора түзу, эмбриондардың дамуы немесе ұлпалардың жіктелуінің болсын негізін құрайды. Әртүрлі клеткаларда гендердің түрлі жиынтықтары транскрипцияланады. . Бұл процестердің әр кезеңінде өзіндік ерекшелігі бар белоктар синтезделеді.
ДНҚ - дан белокқа алып баратын жолда мұндай бақылау кез - келген кезеңде жүзеге аса алады.
Көпжасушалы организмдердің әрбір соматикалық жасушасындағы гендік мәліметтің көп бөлігі бірдей болады. Сондықтан гендік экспрессия бақылануы тиіс . Экспрессия - транскрипция және трансляция арқылы жүзеге асады.
Гендік экспрессияның бақылануы :
1) Оң - белгілі бір регуляцияға жауапты элементтердің әсеріне байланысты, ген экспрессиясының деңгейінің өсуі.
2) Теріс - белгілі бір регуляцияға жауапты элементтердің әсеріне байланысты, ген экспрессиясының деңгейінің төмендеуі.
Негативті бақылану кезіндегі көмектесетін бақылаушы элементті немесе молекуланы негативті бақылаушылар , ал позитивті бақылауға көмектесетіндерді позитивті бақылаушылар деп атайды.
Гендер экспрессиясының реттелуі мынадай деңгейлерде жүруі мүмкін:
- бақылау (геннің транскрипциялану уақыты мен сипаты)
- бақылау (алғашқы транскрипт процессингінің сипаты)
- Транспортдеңгейіндегі бақылау (цитоплазмаға тасымалдауға арналған ядродағы жетілмеген мРНҚ-ны сұрыптау)
- бақылау (рибосомалардағы трансляция үшін қажетті цитоплазмадағы мРНҚ - ны сұрыптау)
- мРНҚ сұрыптау (цитоплазмадағы мРНҚ - ның белгілі бір түрлерін таңдап тұрақтандыру)
Гендер экспрессиясының реттеуші механизмдері туралы мәліметтер микроорганизмдер белоктарының синтезіне қатысатын гендердің активтіліктерінің бақылауын зерттеуде, лямбда фагтың генін, Xenopus гендерін, ашытқылардың шағылысуын қамтамасыз ететін гендерді және эукариоттардың дамуын бақылайтын гендерді зерттеуде алынды. Әртүрлі организмдерде гендер әсерін бақылайтын механизмдерді салыстыру осы механизмдердің алуантүрлілігін көрсетті. Бактерияларда екі механизм белгілі, біріншісі ферменттердің активтілігін, ал екіншісі ферменттердің синтезін бақылайтын механизм (арнайы белоктардың синтезі) . Ферменттердің активтіліктерінің бақылануын (реттелуін) изолейцин аминқышқылы мысалында көруге болады. Яғни изолейцин синтезделу үшін треонин аминқышқылы қажет және ферменттердің қатысуымен бес тізбекті реакциядан өтуі керек. Егер де бактерияның аминқышқылдарын өздігінен синтездей алатын, оның ішінде изолейцинді де, культурасына изолейцин аминқышқылын қосатын болса, онда бактерия клеткалары изолейцин аминқышқылын синтездеуді доғарады. Клеткаға керекті изолейцин бұл кезде тек экзогенді жолмен ғана қамтамасыз етіледі. Бұл құбылыстың механизмі треонин аминқышқылын изолейцинге айналдыратын ферменттің активтілігінің төмендеуімен түсіндіріледі. Қоректік ортада изолейциннің деңгейі төмендеген кезде аталған процесс қайта қалпына келеді.
Бұл құбылыстың бірегейлігі ингибитор (соңғы өнім) және қалыпты субстрат ферменттегі байланысу сайтына таласпайды және олар әртүрлі құрылымға ие болуымен түсіндіріледі. Яғни ферментте екі байланысу сайты болады, оның бірі субстратқа арналған болса, екіншісі ингибиторға арналған. Қалыпты субстрат ферменттің активті сайтына жабысады. Егер де аталған арнайы сайтқа ингибитор байланысатын болса, онда ферментте құрылымдық бұзылыс жүреді (транзиция), нәтижесінде қалыпты субстрат өзінің сайтына байланыспайды, әрі қарай биосинтездің аяқталуын жылдамдататын ферменттің активтілігі тежеледі. Бұл құбылыс аллостерикалық транзиция деген атпен белгілі .
1961ж Жакоб пен Маноның жаңалығы гендер экспрессиясының реттелу механизмдерін анықтаудағы алғашқы қадамы болды. Эукариоттар мен прокариоттар гендерінің экпрессиясы реттеу механизмдерінде кейбір
ұқсастықтар бар, бірақ эукариоттарда генді реттеу жүйесі біршама күрделірек.
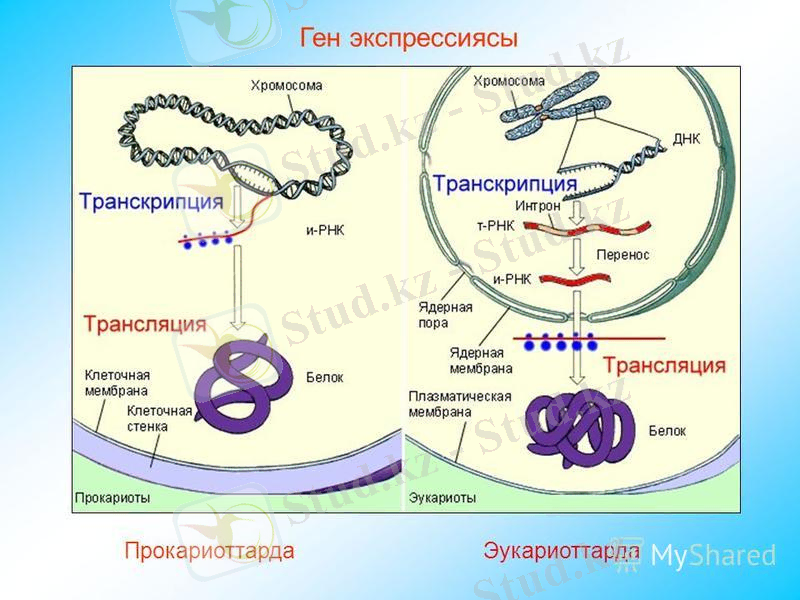
ІІ. 2. ТРАНСКРИПЦИЯ
(лат. transcrіptіo - қайта көшіріп жазу) - генетикалық ақпараттың ДНҚ-дан а-РНҚ-ға көшіріліп жазылуы.
Транскрипция процесі ядрода арнайы фермент - РНҚ - полимеразаның (РНҚ-П) көмегімен ДНҚ және синтезделетін а-РНҚ нуклеотидтерінің комплементарлы жұптасуы жолымен іске асады.
А - РНҚ тізбегінің синтезделуі 5‵ 3‵ бағытында жүреді, яғни жаңа нуклеотидтер синтезделетін а -РНҚ тізбегінің тек 3‵ - ОН ұшына жалғанады. Транскрипция тұтас бір хромосоманың бойында емес, тек ДНҚ - ның бір тізбегінің жеке учаскелерінде жүреді.
ДНҚ молекуласының тізбегі бір - бірінен қызметтері бойынша ажыратылады. Тізбектердің біреуі мағыналы немесе кодтаушы, екіншісі матрицалық (қалып ретінде) тізбек деп аталады. Осы матрицалық тізбек қана транскрипцияланады және РНҚ синтезделуі үшін қызмет атқарады.
Транскрипция процесінің негізгі сипаттамасы:
Барлық ядросы бар жасушаларда ДНҚ - ның транскрипциясы жүріп жатады. Ал, бөліну процесі жүріп жатқан жасушада транскрипция, жасушалық циклдың кез - келген уақытында жүреді, бірақ ДНҚ -ның репликациясы жасуша бөлініп жатқан кезде жүрмейді.
Прокариоттарда ондай шектеу жоқ, олардың жасушалық циклы өте қысқа (20-40 мин. ), сондықтан ДНҚ молекуласының әртүрлі бөлігінде репликация мен транскрипция бір мезгілде жүреді.
Транскрипцияны қамтамасыз ететін процесс
РНҚ - полимеразамен іс - жүзінде асырылады.
Эукариоттарда оның 3 түрі бар:
1. РНҚ - полимераза І - пре - р-РНҚ-ны синтездейді.
2. РНҚ - полимераза ІІ - пре - а-РНҚ-ны синтездейді.
3. РНҚ - полимераза ІІІ - пре - т-РНҚ-ны синтездейді.
Прокариоттарда РНҚ - полимераза 6 суббірліктен тұрады:
1) σ - сигма - транскрипцияның басталуына жауап береді,
2) α, α, β, β‵, ω (омега) - транскрипцияның элонгациясына жауап береді.
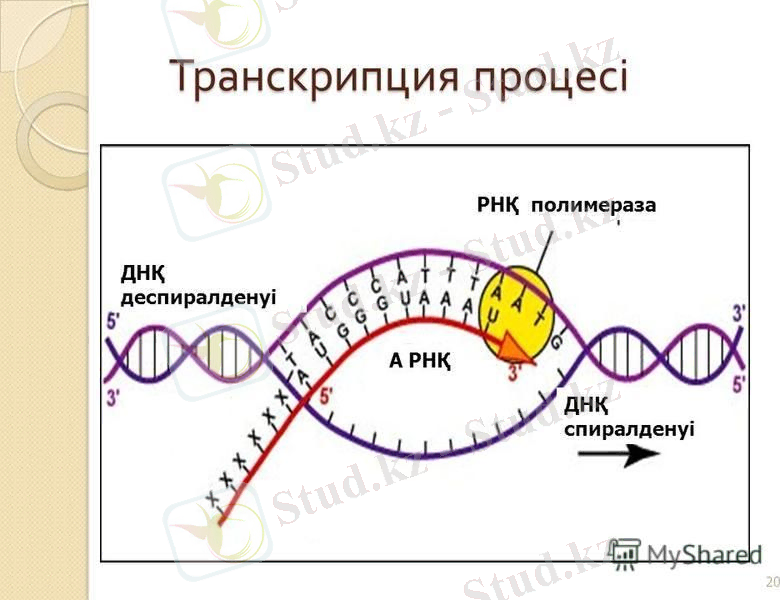
Транскрипция процесі 3 этаптан тұрады:
1. Инициация;
2. Элонгация;
3. Терминация.
Транскрипцияның ең маңызды этапы -
инициация
, онда РНҚ-полимераза промотормен байланысып, нуклеотидтер арасында бірінші байланыс түзіледі.
ІІ. 2. а. Прокариоттардағы транскрипция
Инициация этапы:
Прокариоттарда РНҚ - полимераза матрицалық тізбекті тауып, тікелей промотордың құрамындағы белгілі бір нуклеотидтер жұбының қатарын анықтайды - ол “Прибнов боксы” (5‵ - ТАТААТ - 3‵ қатары) . Оны табатын РНҚ- полимеразаның құрамындағы σ - суббірлігі.
Ол қатарлар транскрипция басталатын нүкте алдындағы нуклеотидтерден «-10» және «-35» ара - қашықтықта орналасқан.
«-10» ауданында ТАТААТ қатары орналасқан, оны “Прибнов боксы немесе домен” деп атайды. (Прибнов 1975г. )
«-35» ауданында ТТГАЦА қатары орналасқан (Travers 1981г. ), оның ДНҚ тізбегінің біреуінде орналасуы, РНҚ- полимеразаға қай тізбектен мәліметтерді көшіру қажет екенін көрсетеді.
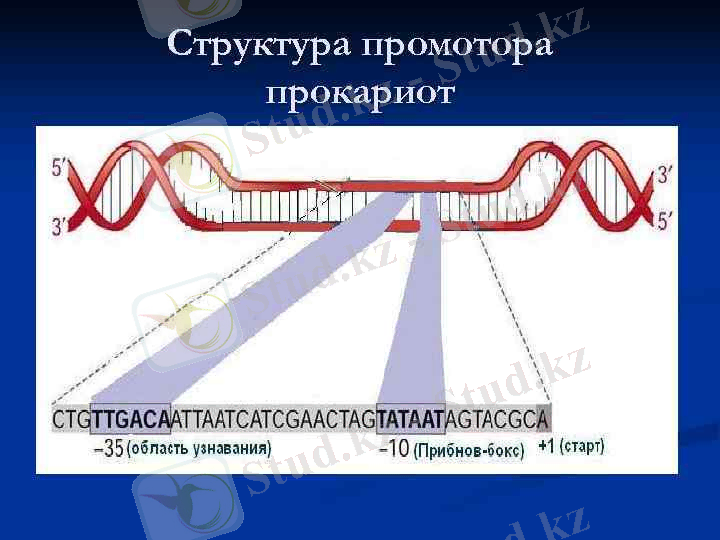
РНҚ полимераза промоторлармен байланысып, ДНҚ- ның қос тізбегін алшақтатады, «-10» ауданындағы нуклеотидтердің айналасында “транскрипциялық шеңбер” пайда болады.
Соған байланысты “шеңбер” аймағындағы ДНҚ-ның матрицалық тізбегінде орналасқан нуклеотидтеріне
рНТФ-тың жұптасуына мүмкіндік туады.
Құрылып жатқан РНҚ тізбегінде бірінші болып пуриндік нуклеотидтер А мен Г орналасып, бірінші мен екінші нуклеотидтер арасында алғашқы фосфорлық байланыс түзіліп, 5‵ 3‵ ұшына қарай бағытталады. Содан кейін
σ - суббірлігі РНҚ- полимеразадан ажырап кетеді.
Инициация процесі кезінде 10 жұп негіздерден тұратын гибридті ДНҚ-РНҚ - олигонуклеотид синтезделеді де, транскрипцияның инициациясы аяқталады.
РНҚ- полимераза ДНҚ тізбектерін тарқатып болған соң
σ - суббірлігі, оның құрамынан ажырап кетеді.
Енді оған тетрамерден (α, α, β, β‵) тұратын РНҚ- полимераза қосылып, элонгация этапы жүре бастайды.
Элонгация
барлық матрицалық процестер сияқты 5‵ 3‵ бағытта жүреді және жаңадан синтезделген РНҚ- молекуласы матрицалық тізбекке комплементарлы болады.
РНҚ тізбегінің ұзаруы РНҚ - полимераза ферментінің көмегімен, рибонуклеозид-трифосфаттардан сол тізбекке рибо- қосу арқылы жүреді.
Транскрипция жылдамдығы бактериялар жасушасында секундына 30-50 нуклеотид.
Прокариоттарда РНҚ молекуласының синтезделген бір бөлігі транскрипция аяқталмай - ақ 5‵ ұшымен рибосомаға байланысып, трансляция жүре бастайды.
Ал, 3‵ ұшында транскрипцияны тоқтататын ДНҚ-ның терминаторлық қатарлары орналасқан.
Терминация
- транскрипцияның соңғы этапы. Терминацияға сигнал болатын геннің соңғы жағында орналасқан Г Ц бай бөліктері.
Прокариоттарда терминацияның 2 типі бар:
1. р-тәуелді, р-факторының қатысуын қажет етеді.
2. р-тәуелді емес.
Прокариоттар мен эукариоттар гендерінің құрылысындағы айырмашылықтар.
Эукариоттар гендері бірінен соң бірі орналасқан ақпараты бар (экзондар) және ақпаратсыз (интрондар) бөліктерден тұрады.
Прокариоттар гендері тек қана ақпараты бар (экзондар) бөліктерден тұрады.
Ақпараттары жоқ бөліктерге интрондар, 5‵ және 3‵ ұштарында орналасқан реттеуші гендер, энхансерлер (транскрипцияны күшейтетіндер), сайленсерлер (транскрипцияны әлсірететіндер) жатады.
Сонымен қатар ақпараты жоқ бөліктерге саттелитті ДНҚ-ғы қайталанатын бөліктер жатады, олар хромосоманың центромерлік және теломерлік аудандарында орналасқан.
Сіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz