Жеке даму биологиясына кіріспе және эмбриологияның даму тарихы

Лекция № 1
Тақырыбы: Кіріспе.
Лекцияның жоспары :
1. Жеке даму биологиясы пәніне жалпы сипаттама
2. Жеке даму биологиясының теориялық және практикалық маңызы.
Лекцияның мақсаты : Студенттерді жеке даму биологиясының зерттеу әдістері және жұмыстарымен, бөлімдері және тарихымен таныстыру.
Лекцияның мазмұны :
1. Жеке даму биологиясы онтогенездің заңдылықтары мен жолдарын зерттейтін ғылым. Қазіргі уақыттағы жеке даму биологиясы бұл эмбриология, цитология, генетика және молекулалық биология салаларының жетістіктерін қамтитын жинақты ғылым. Даму биологиясының негізі эмбриология болып саналады. Жай сөзбен айтқанда даму биологиясының мақсаты: эмбриологиялық, генетикалық, биохимиялық, әдістерді қолдана отырып ұрықтанған жұмыртқадан қандай жолмен күрделі организмнің пайда болатынын түсіндіру болып табылады.
Жеке даму биологиясы гаметогенезді (жыныс клеткаларының дамуын), яғни ұрық пайда болу алдындағы кезеңді, ұрықтық кезеңді және ұрық жарыққа шыққаннан соңғы (постэмбрионалды) кезеңдерді зерттейді. Постэмбрионалды кезең өсу, метаморфоз, жыныссыз көбею, регенерация, соматикалық эмбриогенез сияқты процестерді қамтиды. Сонымен, бұл ғылымның саласы тірі организмдердің дербес дамуы негізіне жататын сан алуан құбылыстарды қарастырады.
2. Эмбриологияның даму тарихындағы бірінші кезең сипаттамалық кезең деп аталады. Ол кезеңде ғалымдар өз зерттеулерінде дамушы организмдегі морфологиялық өзгерістерді суреттеумен ғана шектейтін. Сонымен бірге, алғашқыда ұрықтың түрлі кезеңдеріндегі жалпы құрылысы зерттелген болса, кейінірек зерттеушілердің негізгі назары даму процесіндегі ішкі мүшелер мен олардың нәзік құрылымының пайда болуы мәселелеріне көшкен. Бүгінгі күнде де классикалық сипаттамалық эмбриология өзінің маңызын жойған жоқ, себебі қазіргі заманның электрондық жəне жарық микроскопияның жаңа əдістері қолданылып əр түрлі жануарлардың ұрықтық дамуы жүйелі түрде зерттелуде.
Жануарлар организмдерінің ұрықтық дамуы туралы деректі мəліметтер біртіндеп жиналған сайын, эмбриологияда зерттеудің тəжірибелік əдістерін қолдануға лайықты жағдай жасалады. Сипаттамалық эмбриология əр түрлі ұрықтық даму процесі қалай жəне қандай кезекпен өтетінін зерттесе, эксперименттік эмбриология даму барысындағы əр түрлі процестер мен ұрық бөлшектері арасындағы өзара қарымқатынастарының байланысын анықтауды көздейді. Эксперименттік эмбриологияның негізін қалаушылардың бірі Вильгельм Ру оны «даму механикасы» деп бекер айтпаған. Əр түрлі физикалық, химиялық, ішкі жəне сыртқы факторлардың ұрықтың дамуына тигізетін əсері бүгінгі күні аса зор ықыласпен зерттелуде.
ХХ ғасырдың ортасына қарай дамуды айқындайтын сан алуан себептердің арасында генетикалық фактордың аса маңыздылығы анық болды. Бұл жағдай эмбриогенезді биохимиялық жəне молекулалық биологиялық тұрғыдан зерттеуге үлкен ықылас туғызды, солардың арқасында даму процесінің негізін қалайтын нəзік механизмдер табылды.
Қайсыбір іргелі ғылым болмасын оның қоғамдық пайдасы практикаға қосқан үлесімен есептеледі. Жалпы даму биологиясының, оның ішінде эмбриологияның іс жүзіндегі міндеттері, ең алдымен, жасанды өсіпөнудің жаңа əдістерін табу, гаметалар мен дамудың ерте кезеңіндегі ұрықтарды сақтау əдістерін жетілдіру, эмбриондарды көшіріп енгізу, аса бағалы жануарларды клондау болып табылады. Сонымен қатар, жеке даму биологиясының мақсаты жануарлар мүшелері мен ұлпалардың əдеттегі жəне қатерлік өсуін, жаңаруын, клеткалар мен ұлпа жүйелерінің қартаю мəселелерін зерттеу болып табылады.
Бақылау сұрақтары:
- Жеке даму биологиясы пәннің басқа биологиялық пәндермен тығыз байланысы.
- Жеке даму биологиясының теориялық және практикалық маңызы.
- Жеке даму биологиясының әдістерін суреттеп түсіру, салыстырмалы, эксперименттік, цито-гистолониялық, молекулалы, биологиялық, биохимиялық, иммунологиялық, ген инженериялық.
- Жеке даму биологиясы пәннің медицина, мал өсіру және басқа да салалардағы маңызы.
- Жануарлардың онтогенез кезеңденуі.
Лекция №2
Тақырыбы: Жеке даму биологиясы пәнінің қысқаша даму тарихы
Лекцияның жоспары :
1. Эмбриология, дербес даму биологиясының негізі.
2. Эмбриология көне дәуір және орта ғасырлар кезеңі тарихы.
3. 19-20ғасырлар эмбриология тарихы.
Лекцияның мақсаты Тірі организімнің дамуы туралы /адамдардың/ ғалымдардың көз қарастарымен таныстыру.
Лекцияның мазмұны :
1. Жеке даму биологиясының негізі эмбриология болып табылады. Сондықтан жеке даму биологиясының тарихы эмбриологияның тарихымен тығыз байланысты. Жануарлардың ұрықтық дамуы туралы алғашқы деректер ертедегі өркениет орталықтары - Көне Мысыр, Вавилон, Ассирия, Греция, Үнді мен Қытай елдерінің тарихи құжаттарында кездеседі.
Көне Үнділер (біздің дəуірге дейін бірнеше ғасыр бұрын) ұғымына сəйкес ұрықтың дамуы дененің барлық мүшелерінен алынатын аталық өндіру заты мен (тұқым деген мағлұматқа сай) аналық өндіру заты (бұл менструальды қанымен теңестірілсе керек) қосылуынан басталады. Ұрықтың жынысы ұрықтану кезіндегі “қан” мен “тұқымның” салыстырмалы мөлшеріне байланысты деп саналған. Жалпы айтқанда, осы ұғымдар материалистік көзқарасқа жақын болған. Біздің дəуірге дейінгі (б. д. д. ) VІ - 1 ғғ. үнді медициналық шығармаларында тірі жандардың өзгермейтін тұқым қуалаушы қасиеті бар, сондықтан бала атаанасына ұқсас деген пікір айтылады. Көне Грециядағы жануарлардың эмбриондық дамуы туралы теориялықфилософиялық көзқарастар мен практикалық деректері ерекше назар аудартады. Мысалы, философнатуралист Эмпедокл (б. д. д. 490430 жж. шамасы) ұрық еркек пен ұрғашы ұрықтардың араласу нəтижесінде пайда болады, ал ұрықтың жынысы даму кезеңіндегі температураға байланысты деп санаған. Демокрит (б. д. д. 460370 жж. ) ұрықтың құрамында дененің барлық бөлімдері мен мүшелерінен шыққан бөлшектер барын болжаған. Көне дəуірдің аса көрнекті емшісі Гиппократ (б. д. д. 460377 жж. шамасы) ұрық дененің барлық мүшелерінен шығады деген идеяны қолдаған жəне қандай да болса зақым алған атаанадан дені сау балалармен қатар, ауру балалар тууы мүмкін деп санаған. Ұрық, Гиппократтың көзқарасы бойынша, еркек пен ұрғашы ұрығының араласуынан пайда болады. Ұрықтың айналасында қапшық түзіледі, дем алатын кіндік қалыптасады. Ұрықтың барлық мүшелері бір уақытта құрылады. Яғни, ұрықта барлық органдар түгел, олардың кейінгі даму процесінде жай ғана көлемі ұлғаяды. Гиппократтың бұл ұғымы ХVІІXVІІІ ғасырлардағы көпшілік мақұлдаған преформизм теориясымен үйлеседі. Бұл теория бойынша, қандай да ұрық болсын - ол ересек организмнің кішкентай көшірмесі, даму процесінде ол жай ғана өзгермей (дифференциациясыз) өседі. Дегенмен, Гиппократтың пайымдауынша, дамудың ең алғашқы кезеңінде мүшелер қалыптаса бастайды.
Көне дəуірдің асқан ойшылдарының бірі Аристотельдің (б. д. дейін 384328 жж. ) еңбектерінде биология мəселелері маңызды орын алған. Соның ішінде ол жануарлардың дамуына ерекше назар аударады.
Аристотельдің ұғымы бойынша, ұрық еркек ұрығымен ұрғашының денесінен бөлініп шыққан заттардың қосылуынан пайда болады. Ұрғашы затынан материя (ұрық пайда болу мүмкіндігі) қалыптасады, бірақ бұл мүмкіндікті іске асыру, тек еркек ұрығынан шыққан форманың материяға көрсеткен əсеріне байланысты. Сонымен, еркек ұрықта организмнің мақсатқа лайықты құрылысы мен тіршілік əрекетін түсіндіретін, даму принципі (рух) қаланған.
«Жануарлардың пайда болуы туралы» деген атты трактатында Аристотель жануарлар мен адам эмбриологиясының мəліметтерін талдап қорытады. Ол жануарлардың жыныс айырмашылықтарын, жұмыртқа салушы жəне тірі туушы жануарлардың көбею тəсілдерін, əсіресе адамның көбеюін толық сипаттаған. Тауық эмбрионының дамуындағы арнайы бояусыз, қарусыз көзбен көруге болатын нəрсенің бəрін егжейтегжейіне дейін суреттеген. Ол жыныс тегі, тұқым қуалау, кемтарлықтың пайда болуы, көптұқымдылық жəне постэмбрионалдық даму процесінде белгілер қалыптасуға байланысты сұрақтармен шұғылданған.
Аристотельдің практикалық білімінің деңгейі туралы мынадан түсінуге болады. Ол сүтқоректілерді сипаттаушы ең басты белгілердің ішінде олардың тірі туатынын, төлді сүтпен өсіретінін, ұрықтары жатырға кіндік жəне кейін шу деп аталған мүше арқылы бекінетінін атап көрсеткен. Осы себептен, ол балық тобынан киттерді бөліп алып оларды сүтқоректілер қатарына қосты. Қате деп саналып жүрген Аристотельдің бақылауы: Галеос акуласының жұмыртқасы жатыр қабырғасына сүтқоректі жануарлардың шуына ұқсас мүше арқылы бекінетінін, тек ХІХ ғасырдың ортасында ғана Иоганн Мюллер растады. Аристотель еңбектерінің мəні, ең алдымен, олар болжаудан бақылауға жəне суреттемеге көшуді бейнелейді. Бірақ ерте ұрық пен оның бөлшектерінің нəзік құрылысын терең зерттеуге үлкейткіш оптикалық жүйелер шыққанға дейін көп уақыт бойында мүмкін болмады.
Гиппократқа қарағанда, Аристотель ұрықтың мүшелері біртебірте, бірінен соң бірі біртекті массадан пайда болады, олардың бəрінде алғашында жеке мүшелер болмайды деп санаған. Ақырында, организмнің даму барысында біртіндеп құрылымының күрделенуі туралы Аристотельдің көзқарасы эпигенез теориясының негізін қалады.
2. «Орта ғасырлар» деп аталған V - XV ғғ. кезеңі феодализмнің пайда болуы мен дамуы, сонымен қатар, ғылым, мəдениет, техниканың құлдырауы жəне шіркеу идеологиясының күрт күшеюімен байланысты. Осы кезеңде, табиғат туралы көзқарастар діни қағидалар мен феодалдық қоғам ережелерінің ауыр таңбасын арқалаған еді. Биологиялық білімнің одан əрі кеңеюі мен жүйеленуі XVXVІІІ ғасырларда қайта өрлеу дəуірінде өтті. Биология ғылымының жедел дамуына біріншіден, жаңа техникалық құралдарды (микроскоп, термометр, барометр т. с. ) пайдалану, екіншіден, ғалымдардың көптеген саяхаттарға шығуы себеп болды. XVІXVІІІ ғғ. жануарлар эмбриологиясы саласында да едəуір табысқа қол жетті. Италия ғалымы У. Альдрованди (15221605 жж. ) Аристотельден кейін бірінші рет тауық жұмыртқасының даму сатыларын жүйелі бақылап шығуға тырысты. У. Альдровандидың шəкірті В. Койтер (15341576 жж. ) балапан эмбриогенезін үлкен ұқыптылықпен зерттеген.
Эмбриология тарихында маңызды із қалдырған В. Гарвейдің «Жануарлардың пайда болуы жөнінде» (1651 ж. ) деген кітабы болды. Бұл кітабында ол ұрық даму процесінде мүшелердің жаңадан пайда болуы туралы жəне эмбриогенез жөніндегі кейбір ескі көзқарастарды сынаған (сондайақ, ұрық аталық ұрықтан жəне аналық қаннан пайда болады деген пікір) . Гарвей «тірі жанның бəрі жұмыртқадан дамиды» деген атақты афоризмнің иесі. Ол кездегі, тірі емес заттың өзінен өзі тіріге айналып өсіпөнуі мүмкін деген көзқарастың басымдығын ескерсек, бұл афоризмнің батыл да мағынасының терең екеніне көз жеткіземіз.
1677 жылы С. Гам жəне А. Левенгук адам шəуетіндегі сперматозоидтардың ролін түсінбей, оны ұрық сұйықтығындағы инфузория сияқты паразиттік жəндіктер деп қарастырған.
Бұдан бұрынырақ, 1672 жылы, Р. де Грааф көптеген сүтқоректілердің аналық безіндегі фолликулдарды (ішінде сұйық заты бар, көлемі əр түрлі көпіршіктер) сипаттаған жəне оларды жұмыртқа деп атауды ұсынған, өйткені олар құстардың аналығындағы жұмыртқаға өте ұқсас болған. Ұрықтанғаннан кейін ұрғашы үй қоянының жыныс мүшелерін белгілі бір уақыт сайын зерттеп Грааф аналықтағы көпіршіктердің жарылып босайтынын, ал жатыр түтігінде, саны сол шамадай, олардан едəуір ұсақ жұмыртқалар пайда болатынын көрген. Грааф анағұрлым күрделі, жүздеген клеткадан тұратын құрылымды қателесіп жұмыртқа деп санаған, ол қазір «Грааф көпіршігі» деп аталады, ал шын жұмыртқа осы көпіршіктің ішінде орналасады.
Антуан ван Левенгук (1632-1723 жж. ) өзі жасаған микроскоп көмегімен (үлкейтуі 270 рет) бірқатар омыртқасыз жəне омыртқалы жануарлардың сперматозоидтарын суреттеп, иттің жатыры мен жатыр түтігінде шағылысудан кейін саны аса көп тірі сперматозоидтарды байқаған. Осыған сүйеніп ол жатырдағы ұрық сперматозоидтардан пайда болады деген болжам жасайды.
Қайта өрлеу дəуірінде преформистік жəне эпигенетикалық көзқарастар біржолата қалыптасты. Преформизмді жақтаушылар (Д. Альдрованди, Я. Сваммердам, А. Левенгук, Лейбниц, Боннэ, Галер ж. б. ) ұрықтық даму бұрыннан жұмыртқада (Сваммердам, Валисниери) немесе сперматозоидта (Левенгук, Либеркюн ж. б. ) қалыптасқан нəзік құрылымдардың сапалық жағынан өзгермей тек сандық жағынан өркендеуімен байланысты деп санады (1сурет) .
Анималькулистер (дайын организм спермий ішінде болады деген теорияны жақтаушылар) бағытының ең үздік өкілі Н. Анри - жұмыртқа құрылысы тесігі бар шар тəріздес, ал спермий түрі кішкентай адамшықтарға ұқсас - деп жазған. Адамшықтардың үлкен тобынан санаулысы ғана жұмыртқа ішіне еніп, ішінен жабылып алып, өсе бастайды.
Нағыз преформистер, мысалы Боннэ, «бірінің ішіне бірі салынып қойылған ұрықтар» концепциясын қолдаған. Бұл көзқарас бойынша ұрықтың аналық безінде келесі ұрпақ ұрықтары жатады, ал олардың ішінде өз алдына болашақ ұрпақтардың ұрықтары болады. Сонда Боннэ, Еваның аналық безінде бүкіл болашақ адамзаттың ұрықтары болған деп санады.
Бір ғажабы, сол кездегі преформистік көзқарас табысына микроскопиялық техника кеңінен пайдалана бастауы себеп болды. Микроскопистердің алғашқы зерттеулеріненақ эмбрион құрылысының, тіпті дамудың ерте кезеңінің де орасан зор күрделілігі айқындалды.
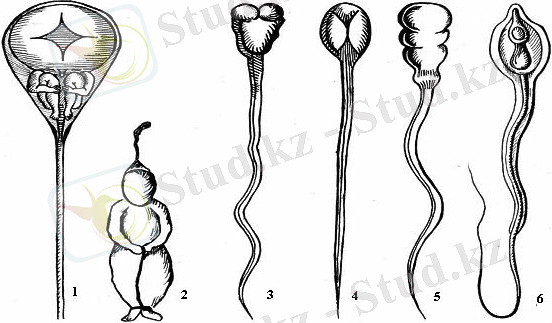
1-сурет. Спермий ежелгі авторлардың көзқарасы бойынша
1 Гартсокер, 2 Далленваций, 3-5 - Левенгук, 6 Пуше
Эпигенетиктер мен преформистер арасындағы айтыста болашақ Ресей академигі Каспар Фридрих Вольфтың (1734-1794 жж. ) «Дүниеге келу теориясы» деген магистрлік диссертациясы аса маңызды роль атқарды. Микроскоппен ұрықтың жеке мүшелерінің түрін жəне олардың пайда болған мерзімін зерттеп, ол мүшелер бір мезгілде емес керісінше бірінен кейін бірі эмбриогенез процесінде дамиды деген қорытындыға келді. Демек, даму процесі эпигенез жолымен жүреді. Мүшелердің дамуы ол жай ғана өсу емес, ол нағыз жаңадан пайда болуының үлгісі. Мысалы, ішек алғашқыда жазық жапырақша түрінде болады да, кейін оралып түтікке айналады.
3. Эмбриологияның биология саласы болып өздігінше қалыптасуынан кейінгі, одан əрі өрлеуі Х. Пандер (1794-1865жж. ), К. Бэр (1792-1876 жж. ), М. Ратке (1793-1860 жж. ) еңбектерімен тығыз байланысты болды.
Пандердің балапан дамуын (1818 ж. ) бақылау нəтижесінің ең маңызды жаңалығы, белгілі сатыда тауық ұрығы үш қабаттан тұратындығы: сыртқы серозды, ішкі - шырышты жəне ортаңғы тамырлы. «Жануарлардың даму тарихы. Бақылаулар мен ойлар» (1 том 1828 ж, ІІ том 1837 ж. шықты) деген классикалық еңбегінде К. Бэр тауық ұрығының дамуын ұсақтүйегіне дейін қадағалады жəне ұрық бейнесі алғашқы біркелкі массадан біртіндеп пайда болатынын, мүшелері кейін қалыптасатынын суреттеді. К. Бэр екі алғашқы анималдық жəне вегетативтік жапырақшаларды ажыратты. Анималдық жапырақша кейін екі қабат береді: терілік жəне бұлшықеттік, ал вегетативтік - тамырлы мен шырышты қабаттарды түзеді. Терілік қабат, өз кезегіне, жамылғыны, нерв жүйесін жəне сезім мүшелерін береді. Бұлшықеттік бұлшықетті жəне сүйекті жасауға негіз болады. Тамырлы - ішперде мен тамырларды, ал шырышты қабат ішек қабырғасын түзеді.
Преформистер мен эпигенетиктер арасындағы қызу таласта К. Бэр көп уақыт бойы бейтарап жол ұстап жүрді, сосын, барлық дəлелдер мен фактілерді пысықтау нəтижесінде, даму преформацияланған эпигенез болады деген тұжырымға тоқтады.
ХІХ ғасырда биологиядағы аса ірі жаңалық клеткалық теория мен эволюциялық ілімнің қалануы екені жалпыға мəлім. Ч. Дарвиннің теориясы тірі табиғат туралы ғылымда төңкеріс жасады. Эволюциялық теория басқа биологиялық ілімдер қатарында эмбриологияға да өз əсерін тигізді. Сонымен қатар, ұрықтық дамуды салыстырмалы зерттеу саласындағы ашылған жаңалықтар эволюция теориясына көп мəн берді, өйткені өздерінің ұйымдасуы бойынша арасы алшақ жануарлар топтарының филогенетикалық туыстығына сенімді дəлелдер келтірді.
Эволюциялық эмбриология ғылымының пайда болуына орыс ғалымдары А. О. Ковалевский (1840-1901 жж. ) мен И. И. Мечников (1845-1916 жж. ) орасан зор үлес қосты.
Сол кездегі зоологтар алдында кейбір типтердің: құрттар, бассүйексіздер, қабықшалылар, иықаяқтылар, губкалар, қылтанжақтылар, ішекқуыстылар, буылтық құрттар, бүйіржүйкелілер, күрекаяқтылар мен бас аяқты моллюскілер, шаянтəрізділер, өрмекшітəрізділер мен бунақденелілердің - шынайы систематикалық орнын, демек, қалған жануарлар əлемімен филогенетикалық қатынастарын табу мəселесі тұрды.
А. О. Ковалевский бірінші болып омыртқасыздардан ұрық жапырақшаларын тапты жəне барлық жануарлардың дамуы бірыңғай жоспармен жүретінін дəлелдеді. А. О. Ковалевский мен И. И. Мечников дəлелдеген ұрық жапырақшаларының гомология идеясы барлық Меtаzоа пайда болуы бірлігінің эмбриологиялық дəлелі болды.
Эмбриология ғылымында суреттеу мен салыстырмалы кезеңдерінен соң ХІХ ғасырдың 7080 жж. эксперименттік бағыт туды. Эмбриологиялық зерттеулерде эксперименттік əдістерді пайдалану нəтижесінде, даму механизмін анықтау жəне эмбриогенез процесіне мақсатты ықпал ету мүмкіндігін зерттеу қажет болды. Жаңа бағыттың қалыптасуы, ең алдымен, неміс ғалымдары Г. Дриш (1867 - 1941 жж. ), В. Гис (1831-1904 жж. ), В. Ру (1850-1924 жж. ), Г. Шпеман (1869-1941 жж. ) есімдерімен байланысты.
Вильгельм Ру «даму механикасы» деп өзі атаған еңбегінде эксперименттік эмбриологияның басты мəселесі себептілік факторларды, дамуды белгілейтін механизмдерді табу деп санаған. Ол даму қатаң детерминацияланған жəне эмбриогенездің барлық кезеңдерінде өсіп келе жатқан ұрықтың барлық бөлшектері өзара тығыз байланысты деп ойлаған.
Көрнекті эмбриолог жəне анатом В. Гис заңды түрде аналитикалық эмбриологияның негізін қалаушы болып есептеледі. Ол бірінші болып, физикахимиялық методтарды қолданып, ұрық дамуының алғашқы кезеңдеріндегі морфогенез процестерінің талдауын жасады. Оның пікірі бойынша, келешек мүшелердің əлі қалыптаспаған бастамалары жұмыртқада немесе əлі дифференциалданбаған ұрықта тəртіппен топталған жəне оларды таңбалап картаға түсіру мүмкін.
Г. Дриш эпигенетикалық көзқарастарды ұстанды. Ол эксперимент арқылы ұрықтың бұзылып барып, дамудың табиғи жолын қайтадан құра алатын қабілетін көрсеткен. Ол теңіз кірпісінің ерте ұрығын қыл тұзақпен екі бластомерге бөліп, олардың əрқайсысынан да толық ұрық дами алатынын көрсеткен. Оны Г. Дриш эмбрионалды регуляция деп атаған.
Эксперименттік эмбриологияға Г. Шпеман да зор үлес қосты. Ол Г. Дриштің экспериментін қайталап, тритонның ерте ұрығын бөлу нəтижесінде бөлінген бөлшектердің кейінгі тағдыры сұр орақ аталған материал қай түрде таратылғанына байланысты екенін көрсетті. В. Ру мен Г. Дриштің зерттеулер бағыттарын жалғастырып, Ганс Шпеман ұрықтың болашақтағы дифференциациясы мен детерминациясы əр түрлі ұрық бөліктерінің өзара қатынасуы жетекші роль атқаратынын көрсетті. Г. Шпеман ұрықтың сұр орақ материалы жоқ бөлігінде білік мүшелер: нерв түтігі, хорда, т. б. түгелдей болмау фактісін талдай келіп, нерв жүйесі презумптивтік (болжалы) хордомезодерма материалының индукциялық əсерінен эктодермадан қалыптасады деген ойын айтты. Осы жорамалды тексеру үшін Г. Шпеман 1924 ж. айдарлы тритонның хордомезодерманың бастамасын қарапайым тритон ұрығының құрсақ энтодермасының астына қондырғанда, қондырылған хордомезодерма шынымен ие клеткасының нерв түтігіне айналуының индукциялағанын көрді. Осы құбылыс алғашқы эмбрионалды индукция деп аталды. Айта кету керек, пішін құру құбылыстарында эмбрионалдық индуктор əсерімен қатар регуляция процестері де байқалған.
Кейінгі, 50-80 жж. С. Тойвонен, Л. Саксен, П. Ньюкуп пен К. Гробстайн ұрық бөліктерінің өзара индукциялық қатынастарын зерттеп, бірқатар екінші индукцияларды тауып ашты.
Орыс ғалымы М. М. Завадовский құстар мен сүтқоректілердің ұрық «бөлшектерінің» өзара əрекеттесу нəтижесінде əр түрлі жыныстық белгілердің пайда болу механизмдерін айқындауға тырысты. М. М. Завадовский алғашқы рет гаметогенез, эмбриогенез, метаморфоз, регенерация жəне басқа морфогенетикалық процестерінің гормондық реттелуін зерттеді.
Мысалы, қораз айдары бас жамылғы ұлпаларынан аталық жыныс гормондары əсерімен өседі. Аталық жыныс безі алынып тасталса айдардың жойылуына апарып соғады, кезегінше, айдарды ампутациялау аталық жыныс безінің гипертрофиясына ұшыратады. Завадовский өсіп келе жатқан организмде тағы да бірқатар «əрекеттестік жүйелерді» эксперимент арқылы тапқан (аналық жыныс безі - жатыр, гипофиз - аналық жыныс безі, т. б. ) .
Орыс эмбриологы Д. П. Филатов омыртқалылардың есту мен көру мүшелерінің дамуын зерттеді. Соның нəтижесінде жаңа микрохирургия əдістерін ұсынды.
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz