Кроссинговер процесінің механизмдері, молекулалық негіздері және дрозофиладағы тәжірибелік зерттеу

Мазмұны
Кіріспе . . . 3
І Кроссинговер . . . 5
- Кроссинговердің механизмдері . . . 11
- Кроссинговердің молекулалық механизмдері . . . 14
- Кроссинговерге әсер ететін факторлар . . . 16
ІІ Тәжірибелік бөлім
2. 1 Дрозофиламен жүргізілген тәжірибеде хромосомалардың айқасуын анықтау . . . 18
Қорытынды . . . 20
Пайдаланылған әдебиеттер
Қосымшалар
Кіріспе
Курстық жұмыстың өзектілігі: Гомологты жұп хромосомалардың бойында аллельді гендердің бір емес, бірнешеуі орналасатыны белгілі. Кейде сол жұп хромосомалар айқасып, нәтижесінде Х тәрізді фигуралар- хиазмалар пайда болады. 1911 жылы Морган ашқан бұл құбылыс хромосомалардың айқасуы немесе кроссинговер деп аталады. Екі жұп хромосомалар айқасудың нәтижесінде бір-бірімен бөлім алмастырады. Басында бір хромосомада орналасқан А мен В гендері әр хромосомаға, одан әрі әр түрлі гаметаларға тарайды. Кроссинговерге ұшыраған хромосомалары бар гаметалар кроссоверлі, ал ондай хромосомалары жоқтары кроссоверлі емес деп аталады. Хромосомалардың айқасу мөлшерін, кроссоверлі особьтардың процентін ұрпақтың жалпы санына шағып есептейді. Айқасудың өлшем бірлігі ретінде оның бір процентке тең мөлшері алынады. Оны кейде морганида деп те атайды. Морган гендер хромосоманың ұзына бойына тізбектеліп орналасады, ал кроссинговердің мөлшері олардың арасындағы салыстырмалы қашықтықты көрсетеді деп есептеді, яғни кроссинговер неғұрлым жиі болса, гендердің бір-бірінен алшақ орналасқандығын, ал кроссинговер сирек болса, олардың жақын орналасқандығын көрсетеді. Мұны Морган дрозофиламен жүргізген тәжірибелерінде дәлелдеді. Ол Х -хромосомада орналасқан денесінің сары түсін (У), көзінің ақ түсін (W) және айыр қанаттылығын (bi) анықтайтын үш түрлі гендер бойынша гетерозиготалы аналық шыбынды гомозиготалы аталық шыбынмен будандастырды. Гомологты хромосомалар бір емес, бірнеше жерден айқасуы мүмкін. Соған байланысты ол сыңар, қос, үш немесе бірнешеу болып келеді. Кроссинговер неғұрлым көп нүктеде жүрсе, соғұрлым гендердің рекомбинациясы (алмасуы) артып отырады. Хромосоманың бір жерінде болатын кроссинговер өзіне жақын орналасқан екінші бір кроссинговерге басымдылық көрсете алады. Ондай құбылыс интерференция деп аталады.
Курстық жұмыстың мақсаты
1. Кроссинговер процесінің жүру механизмін түсіну.
Курстық жұмыстың міндеттері
1. Кроссинговер процесінің жүру механизмін бақылау.
2. Кроссинговерге әсер ететін факторларды зерттеу.
3. Дрозофиладағы хромосомалардың айқасуын бақылау.
І Кроссинговер процесі
Бір хромосомада орналасқан гендердің айқаса алмай, бірігіп тұқым қуалауын гендердің тіркесуі деп атайды. Яғни, олар бір-бірінен тәуелсіз ажырап кетпей, бірге тұқым қуалайды. Алғаш рет бұл құбылысты 1905 жылы У. Бэтсон және Р. Пеннет хош иісті бұршақ өсімдігіне жасаған тәжірибелерінде ашқан болатын. Бұл заңдылықтың теориялық тұжырымдамасын жасаған және терең зерттеген ғалым Т. Морган болды. Кез келген жоғары сатыдағы организмдерде гендердің саны хромосомалардың санынан артық болады. Мысалы, адамның 46 хромосомасында соңғы деректерге сүйенсек, 26 мыңнан 40 мыңға жуық гендер, ал дрозофила шыбынының 4 жұп хромосомасында 7000 ген бар деген болжам бар. Бір жұп хромосомада бірнеше мың ген болады. Бір хромосомада орналасқан гендерді тіркескен гендер деп атайды. Ол гендер тіркесу тобын құрайды. Жасушаның мейоздың бөлінуі кезінде тіркескен гендер бір гаметаға түседі. Т. Морган тіркесу құбылысын анықтау мақсатында зерттеу нысаны ретінде жеміс шыбынын - дрозофила меланогастерді алады. Себебі оның мынадай ерекшеліктері бар. Олар: Дрозофила меланогастер аз уакыт ішінде ұрпақ береді. 8-10 күн ұрпақтарының саны көп, 2-3 аналық, бір аталықтан 500- 600 ұрпақ шыбын алады өзгерген түрлері көп хромосома саны аз. 4 жұп - 8. Денесінің түсі сұр, қалыпты қанатты шыбынды денесінің түсі қара, жетілмеген қанатты аталырымен будандастырады. Бірінші ұрпақ будандарының барлығы денесінің түсі сұр, жетілген қанатты болады. Бірінші ұрпақ будандарын өз-өзімен будандастырғанда, F2 ұрпақта белгілердің фенотипі бойынша ажырауы тәуелсіз тұқым қуалау заңына сәйкес болмай (9:3:3:1), ата-анасының белгілері бойынша 3:1 сан қатынасында ажырайды. Белгілердің 3:1 қатынасында ажырауын, дененің түсін, қанатының пішінін анықтайтын гендер бір жұп хромосомада орналасатынын көрсетеді. Бұл бір хромосомада орналасқан гендердің бірігіп, тіркесіп тұқым қуалайтынын дәлелдейді. Тіркесудің екі түрі болады. Толық тіркесу және толық емес тіркесу. Толык тіркесу - сирек кездесетін құбылыс. Мысалы, денесінің түсі қара, жетілмеген қанатты аналық дрозофила шыбынын, денесінің түсі сұр, жетілген қанатты аталығымен будандастырғанда, ұрпақтарында белгілердің фенотипі бойынша ажырауы 1:1 сан қатынасында немесе 50%-50% болып, бір хромосомада болғандықтан, толық тіркесу байқалады. Мұнда аналық шыбында 4 гаметаның орнына екі типті b+Vg және bVg+, ал аталықта бір типті bVg гаметалар түзіледі, гаметалардағы гендердің қосылу комбинациясы ата-аналық дараларға сәйкес келеді. Сондықтан да ұрпақтары ата-аналық дараларға (денесі кара, жетілген қанат және денесі сұр, жетілмеген қанат) ұқсас болады. Талдаушы шағылыстыруда F 2 -ұрпақтар 50% (қара дене, жетілмеген қанат) және 50% (сұр дене, жетілген канат) болады. Сонда b+ гені Vg+ генімен, ал b гені Vg генімен тіркескен. Бұл гендер бір-бірінен ажырамайтындықтан, гомологті хромосомалардың учаскелерінде алмасу жүрмейді. Жынысына қарамай, бірдей сан қатынасында ата-аналық белгілері бар ұрпақтар түзіледі. Сонымен Т. Морган зерттеген белгілердің тіркес тұқым қуалауы Морганның тіркесу заңы немесе генетиканың төртінші заңы деп аталады. Гомологты жұп хромосомалардың бойында аллельді гендердің бір емес, бірнешеуі орналасатыны белгілі. Кейде сол жұп хромосомалар айқасып, нәтижесінде Х тәрізді фигуралар-хиазмалар пайда болады. 1911 жылы Морган ашқан бұл құбылыс хромосомалардың айқасуы немесе кроссинговер деп аталады. 10-суретте хромосомалардың айқасу схемасы мен оларда болатын гендердің рекомбинациясы көрсетілген. Екі жұп хромосомалар айқасудың нәтижесінде бір-бірімен бөлім алмастырады. Басында бір хромосомада орналасқан А мен В гендері әр хромосомаға, одан әрі әр түрлі гаметаларға тарайды. Кроссинговерге ұшыраған хромосомалары бар гаметалар кроссоверлі, ал ондай хромосомалары жоқтары кроссоверлі емес деп аталады. Хромосомалардың айқасу мөлшерін, кроссоверлі особьтардың процентін ұрпақтың жалпы санына шағып есептейді. Айқасудың өлшем бірлігі ретінде оның бір процентке тең мөлшері алынады. Оны кейде морганида деп те атайды. Мысалы, жүгерінің екі линиясын будандастырғанда, барлығы 1000 дән алынса, оның 36-сы кроссоверлі болған. Сонда айқасудың немесе кроссинговердің мөлшері
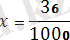
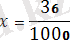 ∙100=3, 6
∙100=3, 6

 .
.
Морган мен оның қызметтестерінің жүргізген зерттеулерінен кейін кроссинговердің әржақты (универсальды) құбылыс екендігі дәлелденді. Ол жүгері, намазшам гүл, тышқан тағы басқалардан да анықталады. Кроссоверлі особьтар санының жалпы оссбьтар санына қатынасын көрсететін кроссинговер мөлшері гендердің ара қашықтығын сипаттайды. Ата-аналық (жұп) хромосомалар арасында екі жақты реципрокты алмасу жүреді, сондықтан кроссоверлі класстар бірге есептелуі керек. Олай болса, жоғарыда дрозофиламен жүргізілген тәжірибенің нәтижесі бойынша екі кроссоверлі кластың әрқайсысы особьтардың 8, 5
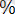
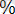 қамтитын болса,
B мен V
гендерінің арасындағы кроссинговердің мөлшері 17
қамтитын болса,
B мен V
гендерінің арасындағы кроссинговердің мөлшері 17

 - ке тең. Морган гендер хромосоманың ұзына бойына тізбектеліп орналасады, ал кроссинговердің мөлшері олардың арасындағы салыстырмалы қашықтықты көрсетеді деп есептеді, яғни кроссинговер неғұрлым жиі болса, гендердің бір-бірінен алшақ орналасқандығын, ал кроссинговер сирек болса, олардың жақын орналасқандығын көрсетеді. Мұны Морган дрозофиламен жүргізіген тәжірибелерінде дәлелдеді. Ол
Х
-хромосомада орналасқан денесінің сары түсін (
У
), көзінің ақ түсін (
W
) және айыр қанаттылығын (
bi
) анықтайтын үш түрлі гендер бойынша гетерозиготалы аналық шыбынды гомозиготалы аталық шыбынмен будандастырды. Нәтижесінде У және W гендерінің рарсындағы кроссинговердің мөлшері 1, 2
- ке тең. Морган гендер хромосоманың ұзына бойына тізбектеліп орналасады, ал кроссинговердің мөлшері олардың арасындағы салыстырмалы қашықтықты көрсетеді деп есептеді, яғни кроссинговер неғұрлым жиі болса, гендердің бір-бірінен алшақ орналасқандығын, ал кроссинговер сирек болса, олардың жақын орналасқандығын көрсетеді. Мұны Морган дрозофиламен жүргізіген тәжірибелерінде дәлелдеді. Ол
Х
-хромосомада орналасқан денесінің сары түсін (
У
), көзінің ақ түсін (
W
) және айыр қанаттылығын (
bi
) анықтайтын үш түрлі гендер бойынша гетерозиготалы аналық шыбынды гомозиготалы аталық шыбынмен будандастырды. Нәтижесінде У және W гендерінің рарсындағы кроссинговердің мөлшері 1, 2

 , W мен bi гендерінің 3, 5
, W мен bi гендерінің 3, 5

 , ал У пен bi гендерінің 4, 7
, ал У пен bi гендерінің 4, 7

 екендігі анықталды. Мұны схема түрінде былай көрсетуге болады:
екендігі анықталды. Мұны схема түрінде былай көрсетуге болады:
1. 2

 3. 5
3. 5


у W -bi
4. 7


Бұл тәжірибенің нәтижесінен кроссинговердің проценті гендердің ара қашықтығына байланысты екендігі көрінеді. Шеткі гендер У пен bi ара қашықтығы екі қашықтықтың У мен W және W мен bi қосындысына тең болғандықтан, гендер хромосоманың бойында бір тізбекпен, кезектесіп орналасады деп тұжырымдау керек. Және әр геннің өзінің сол хромосомада белгілі орны болады, оны локус деп атайды. Гомологты хромосомалар бір емес, бірнеше жерден айқасуы мүмкін. Соған байланысты ол сыңар, қос, үш немесе бірнешеу болып келеді. Кроссинговер неғұрлым көп нүктеде жүрсе, соғұрлым гендердің рекомбинациясы (алмасуы) артып отырады. Мысалы, қос нүктеде болатын айқасуды қарастыралық. Бір хромосомада үш ген бар А - В - С орналасса, айқасу тек А мен В немесе В мен С гендердің арасында ғана емес, қатарынан екі жерде бірдей болады. Сонда тригетерозиготаны талдау жасау үшін будандастырғанда (АаВвСс

 ааввсс) мынадай типті гаметалар береді. [5]
ааввсс) мынадай типті гаметалар береді. [5]
Хромосоманың бір жерінде болатын кроссинговер өзіне жақын орналасқан екінші бір кроссинговерге басымдылық көрсете алады. Ондай құбылыс интерференция деп аталады. Соның салдарынан екі жақты үзілістің проценті нақты жағдайдың процентіне келмей қалады. Бұл әсіресе өте жақын орналасқан гендерден анық байқалады. Гендердің тіркесу мөлшеріне әр түрлі ішкі және сыртқы факторлар да әсер етеді. Мысалы, дрозофиланың кейбір линияларынан алынған будандарда кроссинговер мүлдем болмайды. Бұл көбінесе гомологты хромосоманың біреуінің қайсыбір бөлімінің алмасуына (инверсия) байланысты болады. Кроссинговердің цитологиялық дәлелдемелері: Кроссинговер құбылысы алғашында генетикалық әдіспен дрозофилада анықталып, кейіннен жүгеріде және тетрадалық талдау жүргізу арқылы нейроспрода цитологиялық тұрғыда дәлелденді. Мейоз кезінде байқалатын хромосомалардың айқасуынан хиазмдер пайда болуын кроссинговерді тудыратын механизм деп қарастыруға болады. Бірақ ондай болжам түбегейлі дәлелді қажет етеді. Айқасу процесінде гомологты хромосомалардың бөлімдер алмастыратындығын тұңғыш рет 30- жылдардың бас кезінде цитологиялық тұрғыда дрозофила шыбынында К. Штерн дәлелдеді. Одан кейін Х. Крейтон мен Б. Мак - Клинток дәл сондай заңдылықты жүгері өсімдігінен байқады, Штерннің дрозофиламен жүргізген тәжірибесін қарастырайық. Әдетте екі гомологты хромосома морфологиялық жағынан айнымастай ұқсас болып келеді. Штерн морфологиялық жағынан айырмашылығы бар, яғни толық гомологты емес Х - хромосомаларды алып зерттеді. Бірақ олардың көпшілік бөлігінде гомология (ұқсастық) бар, сондықтан да мейоз кезінде бір-біріне жақындасыпжұптаса алады. Аналықтың Х - хромосомасының біреуі транслокацияның, яғни У - хромосома үзіндісінің айқасуына байланысты Г тәрізді болып өзгерген. Екінші Х - хромосома қалыптыға қарағанда қысқарақ, себебі оның бір бөлігі У - хромосомаға ауысқан. Будандастыру нәтижесінде жоғарыда көрсетілген морфологиялық жағынан әр түрлі екі Х - хромосома және оларда екі ген бойынша гетерозиготалы аналықтан алынған. Оның біреуі көздің пішіні мен көзшелердің санына әсер ететін жартылай доминантты ген В + . Ал В аллелі бар мутанттардың көзі таспа тәрізді болып келеді. Сч гені дрозофиланың көзінің қызыл түсін анықтайда. Г тәрізді Х - хромосомаларда жабайы типке тән В + және Сч + аллельдері, ал қысқалау хромосомаларда мутантты аллельдер В мен Сч болады. Осындай генотипі бар аналық морфологиялық жағынан алғанда қалыпты Сч және В + аллельдері бар Х- хромосомалы аталықпен будандастырылған. Олардың ұрпақтарында кроссоверлі емес хромосомалары бар топ (сч В счВ + , Сч + В СчВ + ) шыбындар пайда болған. Цитологиялық зерттеулер кроссоверлі особьтарда Х - хромосомалардың бөлім алмастыратындығын, соған сәйкес олардың пішінінің өзгеретіндігін көрсетті. Аналық ұрпақтың барлық төрт тобында да атасынан берілген бір-бір қалыпты немесе таяқша тәрізді хромосомадан болған. Кроссоверлі аналық шыбынның кариотипінде кроссинговердің нәтижесінде өзгерген - ұзын таяқша тәрізді немесе қысқа екі иығы бар Х - хромосома болатындығы анықталған. Бұл тәжірибе және осы сияқты жүгерімен жүргізілген тәжірибелердің нәтижесі Морган мен оның шәкірттерінің кроссинговер - гомологты хромосомалардың бөлімдерін алмастыруы және гендер негізінен хромосомаларда шоғырланады деген болжамын дәлелдей түсті. [9]
1. 1. Кроссинговердің механизмдері
Мейоздық кроссинговер: Хромосомалардың айқасуы туралы ғылыми деректердің аз екенінің өзінде - ақ цитологтар мейоздың профазасын зерттеу барысында гомологты хромосомалардың конъюгациялануы (бір - бірімен жанасуы) кезінде Х тәрізді фигуралардың, яғни хиазмдердің пайда болатындығын байқады. 1909 жылы Ф. Янсенс хиазмдер гомологты хромосомалардың бір-бірімен бөлім алмастыруына байланысты деген болжам айтты. Кейіннен бұл 1911 жылы Т. Морган ұсынған хромосомалардың айқасуы жайлы болжамға негіз болды. Мейоздың I профазасының пахинема кезінде гомологты хромосомалардың ұқсас бөлімдері бір - бірімен жақындасып конъюгацияланатыны белгілі. Хромосомалардың ондай күйін бивалент дейді. Әр хромосоманың өзі екі хроматидтен тұрады. Көбінесе сол биваленттің құрамындағы ішкері орналасқан екі хроматид бір - бірімен айқасып, содан барып хиазм пайда болады. Сөйтіп, мейоздың І профазасы кроссинговердің пайда болуының бірден бір сәтті кезеңі болып есептеледі. Жынысты жолмен көбейетін организмдерде кроссинговерді кроссоверлі зиготалардың түзілу жиілігіне қарап анықтайды. Себебі олар кроссоверлі гаметалардың ұрықтануынан пайда болады. Мұны зерттеу үшін объект ретінде зең саңырауқұлағы - нейроспора алынған. Бұл саңырауқұлақтың тіршілік циклінің көпшілігі гаплофазаға сәйкес келеді, ал диплофазасы өте қысқа. Зигота түзілген соң көп кешікпей қайтадан мейоз басталады да, гаплоидты споралары ( спора грек сөзі, тұқым деген мағынаны береді ) бар қалта - аска пайда болады. Сол спораға қарап кроссинговерді оп - оңай анықтайсың. Бұл әдісті тетрадалық әдіс деп те атайды. Сол асканың ішіндегі сегіз спораның төртеуі боялған (), ал қалған төртеуі боялмаған (), олардың қалтаның ішіндегі орналасу реті болып келеді. Егер споралар ААаа ААаа болып орналасса, бұл кроссинговердің болғанын көрсетеді.
Митоздық кроссинговер: Жоғарыда қарастырылған мейоздық кроссинговермен қатар сомалық (дене) клеткаларының бөлінуі кезінде пайда болатын митоздық кроссинговер де кездеседі. 1936 жылы К. Штерн сұр түсті (у + ) денесіндегі қылшығы ұзын (Sn + ) дрозофила шыбынын сары денелі (у) , қылшығы қысқа (Sn) шыбынымен будандастырып, тәжірибе жүргізеді. Сонда олардан алынған дигетерозиготаның екі белгісі де жабайы фенотипке тән болып келеді, бірақта ішінара кейбір шыбындардың денесінде дақтар байқалған. Ондай дақтың тең жартысының түсі сары, қылшығы ұзын, ал қалған бөлігінің түсі сұр, қылшығы қысқа болып шыққан. Мұндай қос дақтың пайда болуын К. Штерн митоздық кроссинговермен түсіндіреді. Шынында, мұндай дақтардың пайда болуы төрт хромотидтер кезеңіндегі гендердің алмасуының нәтижесі болып есептеледі. [7]
Тең емес кроссинговер: Хромосомалар айқасқанда екі гомологты хромосоманың бөлік алмастыратын жері бір нүктеде болады. Соған байланысты екі хромосоманың да айқасқан бөлімдері және олардағы гендердің жиынтығы да сан жағынан бірдей тең келеді. Бірақ кейде, өте сирек жағдайда хромосомалардың айқасуы нүктеде болмай шығады. Соның нәтижесінде хромосомалардың алмасатын бөлімдері және олардағы гендердің саны бір - біріне тең болмайды. Мұндай құбылысты тең емес кроссинговер деп атайды. Мысалы, дрозофиланың жабайы тегіне тән қалыпты көздері хромосомалардың тең айқаспауы салдарынан таспа тәрізді болып өзгерген. [3]
Кроссинговердің механизмі туралы болжамдар: Қазіргі кезде хромосомалардың айқысуының механизмі туралы бірнеше болжам бар, бірақ олардың ешқайсы да гендердің рекомбинациясын және ондағы байқалатын цитологиялық көріністерді жете түсіндіре алмайды. Ф. Янсенс ұсынған, кейіннен С. Дарлингтон ары қарай дамытқан болжам бойынша гомологты хромосмалар бір-біріне жақындасқан кезде олардың жіпшелерінің шиыршықтала бастауына байланысты бивалентке динамикалық тартылыс күші пайда болады. Соның әсерінен төрт хроматидтің біреуі үзіледі. Соған байланысты биваленттің тепе-теңдігі бұзылып, екінші хроматид те үзіледі. Кейін сол үзілген ұштары айқасып, хиазмалар құрайды. Бұл болжам бойынша хиазмалардың түзілуі тікелей кроссиговерге байланысты. К. Сакстың болжамы бойынша, керісінше, алдымен хиазмалар түзіледі де, содан кейін барып кроссинговер пайда болады. Хромосомалар плюстерге ажыраган кезде механикалық күштің әсерінен хиазмалар түзіледі де, сол жерде айқасулар жүреді. Содан соң хиазмалар жойылады. Дж. Беллинг пен Дж. Ледербергтің айтуынша, кроссинговер ДНҚ-ның репликациясы (екі еселенуі) кезінде пайда болады. Сонда бір матрицадан басталған репликация екінші гомологты ДНҚ-ның матрицасына ауысады, соның нәтижесінде генетикалық материал (гендер) рекомбинацияланады. [11]
1. 2 Кроссинговердің молекулалық механизмі
Кроссинговердің молекулалық механзмі жайлы қазіргі көзқарастар негізінен осы ХХ ғасырдың 60 - жылдарында қалыптасты. Соның ішінде әсіресе, хроматидтердің арасындағы рекомбинацияның механизмін түсіндіретін, 1964 жылы Р. Холлидей ұсынған кроссинговердің схемасы ерекше орын алады. Енді рекомбинацияның осы схемасын қарастырайық. Суретте тек екі хроматидтің ғана арасындағы рекомбинация көрсетілген. Қалған екі хроматид соңғы нәтижелерді, яғни тетрадағы ажырауларды қарастырғанда ескеріледі. Схемадағы АВС және авс бір-бірімен тығыз тіркескен таңбалы гендер (маркерлер) . Олардың бағыты бүкіл рекомбинация процесінің барысында қадағаланып отырылады. 12-суретте А бөлігінде хроматид жіпшелерінің үзілген жері бір гомологты нүктелерде болатындығы көрсетілген. Бірінші кезеңде рекомбинацияға кірісетін ДНҚ молекуласы гетеродуплекстер деп аталатын буданды бөлімдер түзеді. Олардың бір тізбегі бір молекуладан, екіншісі басқа молекуладан түзіледі. Бұл жартылай хиазма болып есептеледі. Келесі кезеңде айқасу нүктелеріндегі жіпшелер үзіледі. [8] Сөйтіп, шеткері орналасқан маркерлер бойынша рекомбинантты гетеродуплексті зонада орналасқан екі молекула түзіледі. Дж. Уотсон мен Ф. Криктің айтуынша, мутация дегеніміз - ДНҚ молекуласындағы нуклеотидтердің орналасу ретінің өзгеруі. Ондай жағдайда ДНҚ - ның гетеродуплексті бөлімінде азотты негіздер дұрыс жұптаса алмайды. Ондай өзгерістер арнайы репарация (қайтадан қалыпқа келтіретін ) ферменттері арқылы түзіледі, яғни комплементарлы қалыпқа келеді. Мұндай түзету (коррекция) процесінде матрица рөлін гетеродуплекстің жіпшелері атқарады. Ең соңында жүретін тетрадалық ажырауды осы процестің нәтижесі лдеп қарастыруға болады. Сонымен, жоғарыда айтылған жалпы кроссинговердің механизмін көрсететін жетістіктерге қарамастан әлі де түсініксіз мәселелер көп. Соған байланысты рекомбинацияның молекулалық механизмін зерттеу молекулалық генетиканың басты бір проблемасы болып есептеледі. [9]
1. 3 Кроссинговерге әсер ететін факторлар
... жалғасыСіз бұл жұмысты біздің қосымшамыз арқылы толығымен тегін көре аласыз.
- Іс жүргізу
- Автоматтандыру, Техника
- Алғашқы әскери дайындық
- Астрономия
- Ауыл шаруашылығы
- Банк ісі
- Бизнесті бағалау
- Биология
- Бухгалтерлік іс
- Валеология
- Ветеринария
- География
- Геология, Геофизика, Геодезия
- Дін
- Ет, сүт, шарап өнімдері
- Жалпы тарих
- Жер кадастрі, Жылжымайтын мүлік
- Журналистика
- Информатика
- Кеден ісі
- Маркетинг
- Математика, Геометрия
- Медицина
- Мемлекеттік басқару
- Менеджмент
- Мұнай, Газ
- Мұрағат ісі
- Мәдениеттану
- ОБЖ (Основы безопасности жизнедеятельности)
- Педагогика
- Полиграфия
- Психология
- Салық
- Саясаттану
- Сақтандыру
- Сертификаттау, стандарттау
- Социология, Демография
- Спорт
- Статистика
- Тілтану, Филология
- Тарихи тұлғалар
- Тау-кен ісі
- Транспорт
- Туризм
- Физика
- Философия
- Халықаралық қатынастар
- Химия
- Экология, Қоршаған ортаны қорғау
- Экономика
- Экономикалық география
- Электротехника
- Қазақстан тарихы
- Қаржы
- Құрылыс
- Құқық, Криминалистика
- Әдебиет
- Өнер, музыка
- Өнеркәсіп, Өндіріс
Қазақ тілінде жазылған рефераттар, курстық жұмыстар, дипломдық жұмыстар бойынша біздің қор #1 болып табылады.

Ақпарат
Қосымша
Email: info@stud.kz