Полиморфизм генов репарации ДНК XRCC1 и XRCC3 у жителей бывшего Семипалатинского ядерного полигона

МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ
РЕСПУБЛИКИ КАЗАХСТАН
КАЗАХСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ
ИМЕНИ АЛЬ-ФАРАБИ
Биологический факультет
Кафедра генетики и молекулярной биологии
ВЫПУСКНАЯ РАБОТА
« Изучение полиморфизма генов репарации ДНК (XRCC1 И XRCC3) среди жителей, проживающих на территории бывШего Семипалатинского ядерного полигона»
Исполнитель
Студентка 4 курса Хамдиева О. Х.
Научный руководитель
PhD,
старший преподаватель Алтынова Н. К.
Нормоконтролёр Сатылган И. А.
Допущен к защите
зав. кафедрой,
профессор, д. б. н. Айташева З. Г.
Алматы, 2010
СОДЕРЖАНИЕ
1. 6
1. 7
Индивидуальная радиочувствительность
Минисателлитные мутации
17
19
2. 1. 2
2. 1. 3
2. 1. 4
2. 1. 5
3
3. 1
3. 2
Электрофорез
Метод полимеразной цепной реакции
Рестрикционный анализ
Статистическая обработка результатов
Результаты и обсуждение
Изучение полиморфизма генов репарации
Изучение корреляции между полиморфизмом генов
ДНК-репарации и частотой минисателлитных мутаций
Выводы
Заключение
Cписок использованных источников
ПриЛОЖЕНИЯ
21
22
24
25
27
27
29
32
33
34
37
ВВЕДЕНИЕ
Острота проблемы эффектов малых доз, в очередной раз усиленная последствиями испытаний на Семипалатинском полигоне, кажется, достигла апогея, и находит отражение в статьях дискуссионного характера, содержащих огромный фактический материал. В научной, и в публицистической литературе продолжается острый обмен оценками медицинских последствий влияния проводившихся испытаний на местное население между учеными и специалистами, воспитанными на консервативных основах количественной радиобиологии, и теми, кто пришел в радиационную медицину позже [1] . Вторые имеют желание объективно регистрировать отклонения в здоровье пострадавших, которые возникают в динамике послеаварийных лет. Очевидно, в условиях взаимной критики родится взвешенный взгляд на проблему, и потомки получат объективную и полную оценку того, что мы изучаем сегодня [2] .
Уместно заметить, что почти за 60 прошедших лет существенную эволюцию претерпели оценки медицинских последствий для здоровья пострадавшего населения от атомных бомбардировок японских городов Хиросимы и Нагасаки. По мнению исследователей, распространенность заболеваний, включая рак и лейкемию, связана с воздействием ионизирующего излучения в те годы, когда пациенты были детьми или молодыми людьми [1] .
Данные о биологической эффективности и механизмах действия “малых” доз ионизирующего излучения до настоящего времени остаются противоречивыми, поскольку эффекты “малых” доз ионизирующего излучения оценивали, главным образом, экстраполяцией результатов, полученных при облучении экспериментальных животных относительно высокими дозами. Результаты такой экстраполяции привели к созданию нескольких концепций, объясняющих закономерности возникновения генетических повреждений при воздействии “малых” доз ионизирующего излучения на облучённый организм или клеточные культуры, таких как линейная беспороговая концепция; нелинейная, а также концепция, отстаивающая существование явления гормезиса [2; 3] . Однако данные последних лет свидетельствуют о том, что появление генетических повреждений при воздействии “малых” доз ионизирующего излучения не укладывается в рамки какой-либо одной концепции, т. к. характеризуется взаимодействием сложных внутриклеточных механизмов, к которым относятся механизмы эксцизионной репарации [4], репарации двунитевых разрывов ДНК [5], изменения экспрессии некоторых специфических генов [6] .
Известно, что ионизирующее излучение может вызывать у живых организмов детерминистские (лучевая болезнь, сокращение продолжительности жизни, поражение иммунной системы) и стохастические (канцерогенез, генетические изменения) эффекты, которые зависят от дозы, её мощности и времени, прошедшего с момента облучения. В последние десятилетия большое внимание уделяется оценке темпов мутационного процесса и объёма генетического груза в популяциях человека, подвергшихся воздействию неблагоприятных факторов. Для такой оценки применяются несколько подходов: фенотипический анализ, изучающий частоты наследственных заболеваний, врождённых пороков развития и репродуктивную функцию; цитогенетический анализ (рутинный и FISH), изучающий частоты нестабильных и стабильных хромосомных аберраций, анеуплоидий и сестринских хроматидных обменов в соматических клетках человека [7] ; молекулярно-генетический анализ, позволяющий изучать генные мутации в соматических и половых клетках [8] .
Актуальность проблемы: Знания о закономерностях влияния ионизирующих излучений на геном и индивидуальную радиочувствительность человека можно было бы использовать в целях повышения эффекта лучевой терапии опухолей и противолучевой защиты организма, а также при диагностике индивидуальной предрасположенности людей к различным видам онкологических заболеваний.
Цель исследования: Изучение полиморфизма генов репарации ДНК (XRCC1 и XRCC3) среди жителей, проживающих на территории бывшего Семипалатинского ядерного полигона
Задачи исследования:
1. Изучить распостранённость генотипов (XRCC1 Arg194Trp, XRCC1 Arg399Gln, XRCC3 Thr241Met), связаных с нарушениями в работе систем репарации ДНК среди облучённого населения и в контрольной группе.
2. Сравнить полиморфизм генов репарации (XRCC1, XRCC3) с известными данными о мутациях в минисателлитных локусах среди жителей территорий, прилегающих к СИЯП.
- ОБЗОР ЛИТЕРАТУР
1. 1 Воздействие радиации на организм
К настоящему времени получены фундаментальные данные о реализации эффектов ионизирующего излучения (ИИ) на разных уровнях биологической организации: от молекулярной до организменной. Вместе с тем решение некоторых вопросов как теоретического, так и практического характера до сих пор затруднено в связи с отсутствием полных сведений о закономерностях формирования радиационно-индуцированных эффектов, а также частоте их развития в зависимости от интенсивности воздействия и собственно спектра этих эффектов [9] . Причина сложившейся ситуации может быть обусловлена несколькими обстоятельствами:
- существуют ограничения возможностей применяемых методов исследования - эпидемиологических, клинических, лабораторных и пр. ;
- прямая экстраполяция ситуации, развивающейся при воздействии высокой интенсивности («большие» дозы ИИ), как правило, не совпадает с ожидаемым результатом при воздействии низкой интенсивности («малые» дозы ИИ) ;
- изменения, вызванные воздействием ИИ (особенно при низких уровнях облучения) неспецифичны и схожи с таковыми при развитии типовых патологических процессов и заболеваний. Тем не менее, наличие значительного массива уточненных данных о лицах, подвергавшихся радиационному воздействию, а также обоснованный выбор современных методов исследования и анализа, применяемых в комплексе, позволяют надеяться на получение объективной информации и решение поставленных задач [10] .
Воздействие радиации на организм может быть различным, но почти всегда оно негативно. В результате воздействия ИИ на организм человека в тканях могут происходить сложные физические, химические и биохимические процессы. При попадании радиоактивных веществ внутрь организма поражающее действие оказывают в первую очередь α-источники, и только затем β-источники, т. е. в обратной наружному облучению последовательности. α-частицы, имеющие небольшую плотность ионизации, разрушают слизистую оболочку, которая является слабой защитой внутренних органов по сравнению с наружным кожным покровом. Существует 3 пути поступления радиоактивных веществ в организм: при вдыхании воздуха, загрязненного радиоактивными веществами, через зараженную пищу или воду, через кожу, а также при заражении открытых ран. Наиболее опасен первый путь, поскольку, во-первых, объем легочной вентиляции очень большой, а во-вторых, значение коэффициента усвоения в легких более высокое [11] .
Действие радиации на организм в долгосрочном плане проявляется мутациями, а в краткосрочном - лучевой болезнью различной степени тяжести. ИИ воздействует на клетки организма, вызывая разрушение их белковой структуры. Это приводит к образованию свободных радикалов и других продуктов распада. Помимо смерти самой клетки, ее остатки вызывают общее отравление организма. Хотя клетка и имеет мощные возможности для самовосстановления, при интенсивном облучении не помогают и они [11] .
Сложный патогенез радиационных поражений можно разделить на две фазы:
- фаза первичного действия радиации на молекулярном и клеточном уровне: механизмы поглощения радиации, ионизация атомов и молекул, первичные физико-химические процессы, вызванные непосредственным действием радиации в момент облучения и являющиеся пусковым моментом развития дальнейших патологических процессов.
- фаза развития патологических процессов в организме на молекулярном, органном, системном и организменном уровнях.
Механизмы первичного действия радиации на клетки различны. Приведём некоторые теории и гипотезы, которые, как оказалось, не противоречат, а дополняют друг друга.
Теория прямого действия или теория мишени (Н. В. Тимофеев- Ресовский и др. ) объясняет лучевое поражение клетки как результат прямого попадания γ-кванта или ионизирующей частицы в особо чувствительный объем клетки, в "мишень", удар по которому (прежде всего в ядро клетки) ведет к инактивации, гибели клетки. Эта теория была дополнена теорией непрямого, косвенного действия радиации, в частности на воду, занимающую около 80% массы клетки. При облучении воды образуются ионы и свободные радикалы, которые существуют миллионные доли секунды, и могут оказывать повреждающее действие на структуры клетки [12] .
Кислородный эффект выражается в усилении повреждающего действия радикалов в присутствии кислорода, который взаимодействует с радикалами воды гидратированным электроном, образуя окисляющиеся радикалы. Оказывается, при облучении в присутствии нормального содержания кислорода все клетки и организмы более чувствительны к радиации, и, наоборот, любая гипоксия, недостаток кислорода в момент облучения снижает радиочувствительность в 2-3 раза, т. е. при этом повышается сопротивляемость к облучению [6] .
Была выдвинута сульфгидрильная гипотеза (З. Баб, Я. Траевский, др. ) : окисляющее действие радиации на высокочувствительные SH - группы ферментов и белков, и это может приводить к изменениям и нарушениям биоструктур клетки. Эта гипотеза не оправдалась в полной мере, но в состав многих радиопротекторов входят SH-группы. Были выдвинуты теории образования первичных радиотоксинов: липидных радиотоксинов (осуществляют снижение системы антиоксидантной защиты) в результате действия на ненасыщенные жирные кислоты (Б. Н. Тарусов) и хиноидных, при облучении ароматических соединений (А. М. Кузин) [12] .
Следующий важный вопрос в радиобиологии: поражение каких биоструктур имеет наиболее важное значение в радиационном поражении клетки: поражение ядра или цитоплазмы?
В настоящее время все большее признание получает теория о решающей роли действия радиации на генетические структуры клетки, нарушения функций и структуры ДНК, хромосом, ядра. ДНК - это уникальная структура клетки наиболее чувствительная к облучению, повреждение ее чревато различными трагическими последствиями для всей клетки и даже последующих поколений. При облучении в результате прямого и косвенного действия радиации в ДНК наступают различные структурные нарушения: разрывы водородных связей, нарушения оснований и точечные мутации, одиночные и двойные разрывы цепей ДНК, усиление распада ДНК, нарушение мембранного комплекса ДНК, разрывы хромосом и хромосомные мутации. Одновременно нарушаются ее функции: синтез ДНК, наступает задержка митотических делений, нарушается генетический код, синтез РНК, нарушается обмен веществ и т. д. Эти структурно-метаболические изменения ДНК могут привести к гибели клетки. С другой стороны в то же самое время происходит восстановление ДНК с помощью специальных ферментативных систем, причем легче восстанавливаются одиночные разрывы, труднее - двойные разрывы ДНК [13] .
При облучении нарушаются и структуры цитоплазмы. Высокочувствительны внутриклеточные мембраны и митохондрии, их повреждения приводят к нарушениям метаболизма, окислительного фосфорилирования, недостатку АТФ и внутриклеточной энергии. Нарушения в лизосомном аппарате приводят к высвобождению протеолитических ферментов, которые могут вызвать аутолиз клетки. Однако, поскольку такие структуры в клетке многочисленны и менее чувствительны, эти нарушения легче восстанавливаются [14] .
Степень радиационного поражения клетки зависит от дозы (График 1), вида и мощности излучений, условий среды, содержания кислорода, жизненного цикла клетки.
В зависимости от нарушений ДНК и цитоплазмы различают следующие эффекты действия радиации на клетки: интерфазная гибель клеток вскоре после облучения; репродуктивная гибель в процессе митоза или через несколько клеточных делений; в некоторых клетках происходит репарация сублетальных повреждений; появление мутантных клеток с генетическими нарушениями или с онкологическими последствиями.
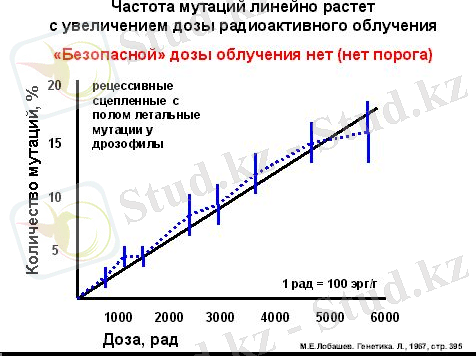
График 1. Зависимость частоты мутации от дозы радиоактивного облучения
Характерной чертой лучевой болезни, отличающейся от многих других заболеваний, является поражение генетического аппарата клетки (ДНК, хромосом), нарушение размножения клеток, массовая гибель радиочувствительных клеток, опустошение радиочувствительных тканей и систем, прежде всего кроветворной системы и слизистой кишечника. Наиболее чувствительны к радиации интенсивно делящиеся клетки, поскольку если в момент деления будет разрушена одна клетка, то погибнут обе. Это ведет к истощению запаса этого типа клеток (если клетки интенсивно делятся, то они примерно с такой же скоростью и умирают) [15] .
Более всего страдают ткани костного мозга и лимфатической системы: эритроциты и лейкоциты постоянно обновляются в организме. Так же чувствительны клетки желудочно-кишечного тракта, клетки волосяного фолликула. Менее всего чувствительны к радиации неделящиеся клетки нервной системы. Из этого следует, дети и подростки более восприимчивы к радиации, чем взрослые, а наиболее чувствителен - эмбрион в утробе.
Эффекты воздействия радиации на человека обычно делятся на две категории (Таблица 1) :
- Соматические - возникающие в организме человека, который подвергался облучению.
- Генетические - связанные с повреждением генетического аппарата и проявляющиеся в следующем или последующих поколениях: дети, внуки и более отдаленные потомки человека, подвергшегося облучению [16] .
Таблица 1.
Радиационные эффекты облучения человека
1. 2 Радиационно-индуцированные хромосомные аберрации
При воздействии радиации на любой живой организм главной мишенью ее воздействия является генетический материал клетки. При этом чувствительность этой мишени превышает чувствительность других биологических мишеней (белков, мембран, надмолекулярных структур в десятки раз) . Причиной радиационной гибели многоклеточных организмов является гибель фракции наиболее чувствительных и незаменимых для жизнедеятельности организма клеток, которые, в свою очередь, гибнут из-за поражения их генетического материала [12] .
Генетический материал всех клеток представлен молекулами ДНК. Огромные полимерные нити ДНК имеют строго определенную первичную структуру, которая должна поддерживаться в неизменном виде в течение многих поколений. Однако это условие при физиологических температурах по чисто термодинамическим причинам не может быть выполнено. Например, в течение часа в ДНК каждой клетки человека возникает несколько десятков тысяч повреждений, которые, как правило, несовместимы с жизнеспособностью клетки. Частота различных типов повреждений коррелирует с прочностью химических связей, разрыв которых является причиной их возникновения.
Спонтанные повреждения ДНК встречаются довольно часто, такие события имеют место в каждой клетке. Для устранения последствий подобных повреждений имеется специальные репарационные механизмы (например, ошибочный участок ДНК вырезается и на этом месте восстанавливается исходный) . Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений. Мутации, возникающие в генах белков, ответственных за репарацию, могут приводить к многократному повышению (мутаторный эффект) или понижению (антимутаторный эффект) частоты мутирования других генов. Так, мутации генов многих ферментов системы эксцизионной репарации приводят к резкому повышению частоты соматических мутаций у человека, а это, в свою очередь, приводит к развитию пигментной ксеродермы и злокачественных опухолей покровов [17] .
При хромосомных мутациях происходят как изменение числа отдельных хромосом в геноме (анеуплоидия), так и крупные перестройки структуры отдельных хромосом. Последние получили название хромосомных аберраций. В этом случае наблюдаются:
1. Делеция
а) интерстициальная - выпадение участка хромосомы в средней ее части б) дефишенская - потеря концевых участков хромосомы;
в) появление кольцквых хромосом
2. Дупликация - двух- или многократное повторение генов, локализованных в определенном участке хромосомы
3. Изохромосомы - плечи одной хромосомы разрываются в центромерном районе; при делении клетки образуются хромосомы с равными плечами (метоцентрик), при этом количество генов удваивается (Рисунок 1(б) ) .
4. Инверсия - поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной;
а) парацентрическая (Рисунок 2(5) ) .
б) перицентрическая (Рисунок 2(4) ) .
5. Транслокация - изменение положения какого-либо участка хромосомы в хромосомном наборе, т. е. перенос генетического материала от одной хромосомы к другой.
а) реципрокная - наиболее распространенная, при которых происходит обмен участками между двумя негомологичными хромосомами.
б) нерецепрокная - участок хромосомы может изменить свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую.
в) робертсоновской транслокация - чаще всего вступают акроцентрики (Рисунок 1(а) ) [18] .
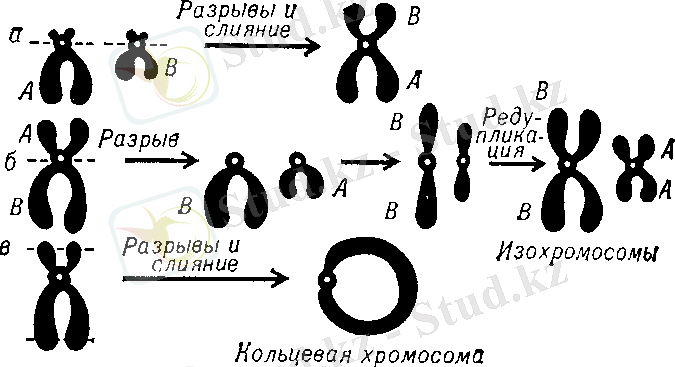
Рисунок 1. Схема образования робертсоновской транслокации (а), изохромосом (б) и кольцевой хромосомы (в)
Примечание: A и В - плечи хромосом

Рисунок 2. Виды внутрихромосомных перестроек
Примечание: 1 - исходная пара гомологичных хромосом; 2 - потеря участка DEFH хромосомы (делеция) ; 3 - удвоение участка С в хромосоме (дупликация) ; 4 - инверсия участка BCD в хромосоме; 5 - инверсия участка DE в хромосоме.
1. 3 Генные мутации
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точковых мутациях (рисунок 3) . Поскольку в состав ДНК входят азотистые основания только двух типов - пурины и пиримидины, все точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот) .
Возможны четыре генетических последствия точковых мутаций:
1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида),
2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация),
3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация) . В генетическом коде имеются три бессмысленных кодона: амбер - UAG, охр - UAA и опал - UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов - например, амбер-мутация),
4) обратная замена (стоп-кодона на смысловой кодон) [19] .
По влиянию на экспрессию генов мутации разделяют на две категории: мутации типа замен пар оснований и типа сдвига рамки считывания (frameshift) . Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трем, что связано с триплетностью генетического кода.
... продолжениеВы можете абсолютно на бесплатной основе полностью просмотреть эту работу через наше приложение.
- Информатика
- Банковское дело
- Оценка бизнеса
- Бухгалтерское дело
- Валеология
- География
- Геология, Геофизика, Геодезия
- Религия
- Общая история
- Журналистика
- Таможенное дело
- История Казахстана
- Финансы
- Законодательство и Право, Криминалистика
- Маркетинг
- Культурология
- Медицина
- Менеджмент
- Нефть, Газ
- Искуство, музыка
- Педагогика
- Психология
- Страхование
- Налоги
- Политология
- Сертификация, стандартизация
- Социология, Демография
- Статистика
- Туризм
- Физика
- Философия
- Химия
- Делопроизводсто
- Экология, Охрана природы, Природопользование
- Экономика
- Литература
- Биология
- Мясо, молочно, вино-водочные продукты
- Земельный кадастр, Недвижимость
- Математика, Геометрия
- Государственное управление
- Архивное дело
- Полиграфия
- Горное дело
- Языковедение, Филология
- Исторические личности
- Автоматизация, Техника
- Экономическая география
- Международные отношения
- ОБЖ (Основы безопасности жизнедеятельности), Защита труда
