Морфометрический и ядерный анализ жестких поляризованных клеток опушения листовой пластинки пшеницы и оценка перспективных линий

Казахский Национальный Университет им. Аль-Фараби
Биологический факультет
Кафедра генетики и молекулярной биологии
Дипломная работа
Изучение жестких клеток опушения пшеницы
Алматы 2010
РЕФЕРАТ
Дипломная работа 37 с., рис. 11, использованных источников 50.
Ключевые слова: ПШЕНИЦА, ЖЕСТКОЕ И МЯГКОЕ ОПУШЕНИЕ ЛИСТЬЕВ, ТРИХОМЫ, ЯДРО.
Объекты исследования: Казахстанская-126 (контроль без опушения), Саратовская-29 и Генотроф-1 (контроли мягкого опушения), Велютинум-2401 и Лютесценс-19001 (константные формы - носители жесткого опушения) .
Цель работы - изучение перспективных линий пшеницы с высокой плотностью жестких поляризованных клеток листовой пластинки.
Задачи:
- провести сравнительный морфометрический анализ клеток жесткого и мягкого опушения;
- изучить закономерности развития клеток жесткого опушения листовой пластинки при онтогенезе пшеницы;
- изучить серию перспективных линий пшеницы с высокой плотностью жестких поляризованных клеток листовой пластинки;
- провести сравнительный морфометрический анализ ядер из клеток жесткого и мягкого опушения.
СОДЕРЖАНИЕ
ВВЕДЕНИЕ 4
1 ЛИТЕРАТУРНЫЙ ОБЗОР 5
1. 1 Морфологические особенности клеток опушения листовой платинки
высших растений 5
1. 2 Типы опушения 8
1. 3 Характеристика развития клеток опушения ( трихом ) 9
1. 4 Функциональное значение трихом 10
1. 5 Опушение листьев как фактор устойчивости к листовредителю -
пьявице 17
1. 6 Значение опушения листьев для засухоустойчивости растений 21
2 МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ 22
2. 1 Приготовление ацетокармина 22
2. 2 Характеристика исследуемых сортов и линий 24
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ 24
3. 1 Морфометрический анализ клеток жесткого и мягкого
опушения пшеницы 24
3. 2 Физиологические особенности поляризованных клеток
листовой пластинки пшеницы 27
3. 3 Параметрический анализ ядер из клеток мягкого и
жесткого опушения 29
ЗАКЛЮЧЕНИЕ 33
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ 34
ВВЕДЕНИЕ
Растение использует разнообразные способы защиты от засухи. Набор определенных морфологических признаков способствует тому, что злаки легче переносят данный, абиотический фактор стресса. Особое значение в защите листа от перегревания и интенсивной солнечной радиации имеет опушение. Большинство сортов, районированных в Поволжье, Западной Сибири и Казахстане в этой связи имеют густое опушение [1] . Наличие жесткого опушения листовой платинки снижает температуру листа, способствует повышению устойчивости к засухе, а также устойчивости к повреждениям вследствие суховеев, пыльных бурь и нашествий листогрызущих вредителей.
Опушение листовой пластинки у молодых растений - это визуально-различимый, маркерный признак, который выполняет защитную роль. В результате реализации данного признака снижается повреждаемость всходов пшеницы при нашествии шведской мухи ( Lajdelphax striatella ), а в фазах трубкования и колошения - пьявицы ( Oulema melanpous ) . Этот вредитель уничтожает паренхиму листьев, не повреждая клеток эпидермиса. Пьявица является наиболее активным вредителем на Юге и Юго-Востоке Казахстана. В отдельные годы этот листоед поражает посевы так, что это ведет к снижению урожайности пшеницы на 25 % [2] .
Актуальность темы. Известно, что клетки опушения являются производными эпидермиса и выполняют ряд центральных функций в растениях [3] . Среди таких ключевых функций: водосбережение, защита от вредителей и патогенов, а также секреция вторичных метаболитов. В последние годы в странах СНГ и дальнего зарубежья можно наблюдать подъем исследовательского и селекционного интереса к новым формам пшеницы с повышенной плотностью опушения. Это объясняется возрастающим комплексом проблем, а именно: 1) усилением действия внешних стрессовых факторов и необходимостью эффективной регуляции продуктивности; 2) истощением почвенных ресурсов и необходимостью поддержания стабильных и возрастающих урожаев; 3) учащением явлений сцепленного генетического загрязнения при гибридизации пшеницы и необходимостью устранения случайного материала с помощью контроля над определенным фенотипическим признаком.
Цель работы - изучение перспективных линий пшеницы с высокой плотностью жестких поляризованных клеток листовой пластинки.
Задачи:
- провести сравнительный анализ морфологии клеток жесткого и мягкого опушения пшеницы;
- изучить наиболее перспективные линии пшеницы с высокой плотностью жестких поляризованных клеток листовой пластинки;
- провести анализ размеров и локализации ядер в клетках жесткого и мягкого опушения;
1 ЛИТЕРАТУРНЫЙ ОБЗОР
1. 1 Морфологические особенности клеток опушения листовой пластинки высших растений
Трудно назвать другую черту строения растительного организма, которая характеризовалась бы таким же разнообразием форм, как поляризованные клетки опушения, или трихомы . Понятно в связи с этим стремление морфологов создать классификацию трихом, свести все огромное их многобразие в более или менее обозримый вид.
По современным данным, имеется 21 канонический тип опушения листовой пластинки. При этом классификация типа опушения зависит от формы, цвета и распределения трихом, а так же их индивидуальной структурной организации [4] .
К числу наиболее распространенных классификации оносится классификация Солередера. Солередер, как и многие другие авторы, делит все типы трихом на кроющие и железистые. Железистые выделяют секрет, тогда как кроющие не обнаруживают секреторной активности.
Кроющие трихомы подразделяются на несколько групп. Первая группа - простые кроющие волоски: трихомы, состоящие из одной клетки, или несколько клеток, расположенных в один ряд. Эта группа включает много различных типов трихом. Из них наиболее просто организованы папиллы, или сосочки. Они образуются в результате выпячивания наружной оболочки клетки эпидермиса, апикальная часть которого несколько загруглена. Обычно из этой эпидермальной клетки образуется один сосочек, но в некоторых случаях, например у эпидермиса листа манника плавающего, одна клетка может иметь несколько выростов. Чаще всего папиллы встречаются на частях цветка. Довольно близки по своей организации к папиллам одноклеточные пузырчатые трихомы. Они также образуются в результате выпячивания наружной стенки протодермальной клетки, но возникает не сосочек, а сферическая структура в вид пузырька. Нередко одноклеточные волоски достигают значительны больших размеров, и их длина во много раз превышает диаметр. Такие трихомы встречаются в трубке венчика вероник.
Особо выделяет Солередер специалные одноклеточные простые трихомы. К ним он относит так называемые цепляющиеся трихомы, апикальная часть которых загнута в виде крючка, Т-образные волоски, возникающие в результате раздвоения верхней части клетки волоска на две ветви. В тех случаях, когда формируется только одна такая ветвь, волосок приобретает форму буквы Г. Иногда клетка волоска, наоборот, дает несколько выростов, направленных в разные стороны, образуя звездчатый волосок. Сюда же относятся и одноклеточные пельтатные волоски, которые в отечественной литературе обычно называются щитовидными. Они формируется вследствие разрастания клетки волоска в плоскостьи, паралельной поверхности эпидермиса.
Многоклеточные трихомы, состоящие из клеток, расположенных в один ряд в типичном случае образованы морфологически сходными клетками. Однако иногда терминальные клетки существенно отличаются по своему строению от остальных. Так, у Atriplex они приобретают сферическую форму. Они иногда терминальные клетки существенно отличаются. Они могут быть щитовидными, звездчатыми. Т-образными, в виде крючка или достигать значительно больших размеров по сравнению с базальными. Иногда все клетки волоска претерпевают изменения. В частности, у Rhinanthus major каждая клетка волоска образует вырост, который, однако, не отделяется перегородкой.
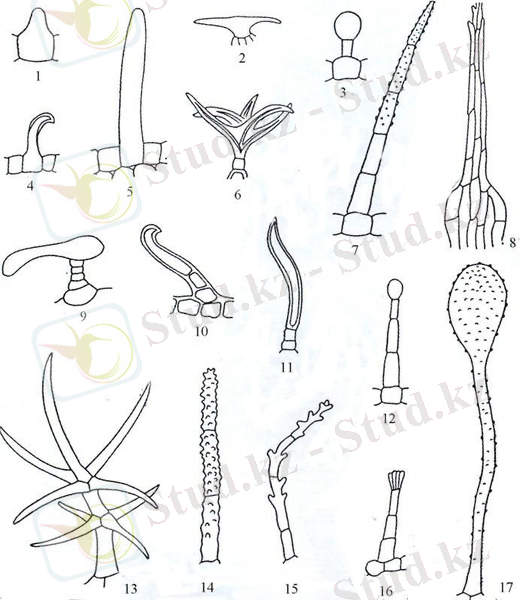
Рисунок 1 - Трихомы растений
1- Viola tricolor ; 2- Humulus lupulus L. ; 3- Atriplex leucoclada Boiss. ; 4- Broussonetia papyrifera Vent; 5- Veronica opaca Fries; 6- Eurotia cerotoides (L. ) G. A. Mey. ; 7- Melampurum nemorosum L. ; 8- Papaver sp. ; 9- Chenopodium botrys L. ; 10- Aristolochia tomentosa Sims. ; 11- Casuarina equisetifolia L. ; 12- Digitalis ambigua Murr. ; 13- Verbacum thapsiforne Schrad. ; 14- Melampurum nemorosum L. ; 15- Rhinanthus major L. ; 16- Antirrhinum majus L. ; 17- Verbacum phlomoides .
К первой группе следует отнести и весьма своеобразные трихомы, встречающиеся на частях цветка, которые были детально описаны для норичниковых. Они довольно крупные (около 0, 7 мм) и отличаются характерным строением оболочек - очень тонкие стенки их клеток образует многочисленные выпячивания, становяськак бы гофированными.
Ко второй группе Солередер относит щитовидные, звездчатые и канделябровые волоски. В отличие от аналогичных типов трихом первой группы - щитовидная, звездчатая части или отдельные мутовки этих трихом образованы не одной, а двумя или большим числом клеток. Щитовидные волоски состоят из дисковидной многоклеточной пластинки и ножки. Пластинка чаще всего образованы одним слоем клеток, длинные оси которых могут располагаться радиально или имеют иную ориентацию. Ножка может быть одноклеточной или состоять из комплекса клеток.
У звездчатых волосков составляющие их клетки разрастаются в виде расходящихся лучей, которые имеют различные ориентацию. У волосков однихрастений они расположены в плоскости, паралельной поверхности эпидермиса, у других расходятся под некоторым углом. Канделябровые волоски в русской литературе называют ветвистыми, так как составляющие их клетки разветвляются, очень часто мутовчато. Типичным примером таких волосков являются кроющие волоски Verbascum.
Третья группа включает волоски, состоящие из несколькмих рядов клеток. Клетки, расположенные в верхней части волоска, часто образуют папиллы. Эти волоски очень сильно различаются по диаметру и длине и нередко достигают значительной величины, образуя густое войлочное опушение. Иногда они ветвятся.
Железистые трихомы подразделяются на две группы. Первая группа включает мелкие волоски простого строения, ко второй группе относятся гораздо более сложные. Границы между этими группами, по словам самого Солередер, весьма условны.
Первая группа разбита на подгруппы 1а и 1б.
Подгруппа 1а включает в себя одноклеточные и многоклеточные железистые волоски. Последние крайне разнообразны по своей форме. Наиболее простые состоят из одного ряда морфологически сходных клеток. Другие имеют «головку» и «ножку». Ножка может быть длинной или короткой, одноклеточной и многоклеточной. Головки также весьма разнообразны по форме и числу составляющих их клеток. При этом многоклеточные головки бывают разделены или периклинальными перегородками или периклинальными и антиклинальными вместе. Иногда перегородки, делящие головку, не имеют определенной ориентации. Среди этой подгруппы Солередер выделяет также щитовидные, сферические или имеющие иную форму железки.
К подгруппе 1б относятся многоклеточные головчатые волоски, головки которых разделены только антиклинальными перегородками.
Во вторую группы входят трихомы наибольших размеров и более сложной организации. У них иногда имеются даже устьица.
Особо выделяют Солередер железистые волоски нектарников, насекомоядных растений, известковые и солевые железки.
Волоски различают не только по их строению, но и по продолжительности существования. В этом отношении они подразделяются на опадающие и неопадающие. Неопадающие волоски сохраняются на протяжении всего периода жизни несущего их органа. При этом протопласт клеток неопадающих волосков нередко довольно рано отмирает и остаются лишь их клеточные оболчки, т. Е. волоски мертвыею Уопадающих волосков с отмиранием протопласта волосок сбрасывается. Отчленение волосков от тела растения обычно соправождается весьма существенными изменениями в структуре клеточных стенок эпидермальных клеток, примыкающих к волоску. Обычно они кутинизируется. У Digitalis ambigua кутинизация в первую очередь распространяется на срединную пластинку и начинается еще до того момента, когда наступает полное отмирание пропласта волоска.
Волоски различаются и по положению относительно поверхности несущего их органа. Одни расположены непосредственно на эпидермисе и называются сидячими. Другие находятся на своебразных выростах.
Выделяют также акроскопные и базископные волоски. Акроскопные более или менее прижаты к поверхности и их вершина ориентирована в направлении роста органа, на котором они располагаются. Базископные волоски направлены противоположно.
Те или иные типы трихом очень бывают присущи определенным группам растений. Поэтому они широко используется в качестве одного из систематических признаков. Так например, у Papilionaceae и Asteraceaeпочти все кроющие волоски - многоклеточные однорядные, тогда как у Cruciferae встречаются лишь одноклеточные волоски. У видов Scrophulariaceae головки железистых трихом делятся вертикальными перегородками, тогда как у Solanaceae- как вертикальными, так и горизонтальными [5] .
1. 2 Типы опушения
В зависимости от строения трихом, их размеров, числа на единицу площади, ориентации и т. п. внешний вид поверхности того или иного органа может быть различным. В связи с этим выделяют определенные типы опушения. В частности, густое опушение из ветвистых волосков несколько напоминает войлок и названо войлочным типом опушения. Наиболее типичное войлочное опушение у коровяка. Многочисленные папиллы придают поверхности бархатистый оттенок. Такое опушение чаще всего встречается у лепестков и называется бархатистым.
Длинные, несколько извилистые волоски, не имеющие определенной ориентации, образуют мохнатое опушение. Мохнатое опушение имеет, например, Begonia rex Putz.
Выделяют шелковистое опушение, состоящее из тонких прямых волосков, обычно ориентированных в одном направлении. Такие волоски придают поверхности листа шелковистый блеск ( Fagus orietalis Lipsky) . У Senecio cineraria DC. опушение образовано очень тонкими, длинными, переплетающимися между собой волосками- паутинистый тип.
Различают также пушистое, волосистое, шерститое, коротковолосое, жестковолосое, шершавое, щетинистое, чешуйчатое и др. типы опушения. Однако хорошо разработанной классификации типов опушения до настоящего времени не существует, поэтому нередко при описании опушения авторы основываются лишь на субъективном восприятии и часто опушениям, состоящим из однотипных волосков, дают разные названия или, наоборот, опушения, значительно различающиеся по структуре составляющих их волосков, относят к одному и тому же типу [6] .
1. 3 Характеристика развития клеток опушения ( трихом )
Одной из наиболее характерных особенностей трихом является их чрезвычайно ранняя дифференциация. Нередко трихомы начинают закладываться на только что образовавшихся примордиях, а иногда - даже на конусе наростания [7] . Заложение трихомы начинается с того, что одна из клеток протодермы, имеющая более крупное ядро по сравнению с остальными, увеличивается в размерах, разрастаясь главным образом в радиальном направлении. При образовании многоклеточного волоска, состоящего из одного ряда клеток, разросшаяся клетка протедермы делится в тангентальном направлении. В результате этого деления образуется одноклеточный зачаток. Он быстро разрастается и делится все новыми и новыми тангентальными перегородками. При этом у некоторых растений наблюдается акропетальная последовательность делений, т. е. все время делится лишь апикальная клетка волоска ( Artemisia ), у других же видов (Helianthus) деления протекают в базальной клетке. Такая последовательность заложения называется базипетальной.
В случае образования папилл дифференциация как бы приостанавливается на самых ранних этапах развития трихом, после выпячивания наружной стенки клетки протодермы. Аналогичная картина наблюдается и при заложении одноклеточных волосков, которые не отделены тангентальной перегородкой от трихоцита, т. е. клетки, из которой образуется волосок.
При образовании головчатых волосков дифференциация ножки обычно происходит таким же образом, как и многоклеточного волоска, состоящего из одного ряда клеток. Головка же может образовываться как путем тангентальных, так и путемантиклинальных делений, а иногда деления протекают без определенного порядка. В тех случаях когда головка одноклеточная, апикальная клетка разрастается, принимая округлую форму.
Дифференциация щитовидных трихом начинается, как правило, делением трихоцита в антиклинальном направлении. Антиклинальные деления преобладают и при дальнейшей дифференциации трихом этого типа. Таковы наиболее часто встречающиеся типы формирования трихом.
Как показали исследования, микрофибриллы наружных слоев первичной оболочки растущего волоска располагаются в направлении, близком к аксиальному, тогда как микрофибриллы внутренних слоев ориентированы поперек длинной оси волоска. При этом в наружных слоях расположение микрофибрилл более рыхлое по сравнению с внутренними [8] . На основании этих данных Релофсен и Хаувинк выдвинули оригрнальное представление о росте оболочке. Микрофибриллы откладываются на внутрепнюю поверхность оболочки в поперечном по отношению к длинной оси волоска направлении. Затем в процесс растяжения клетки они пассивно изменяют свою ориентацию, вытягиваясь примерно в более или менее аксиальном направлении. И поскольку новые микрофибриллы в это слой не внедряются, он становится более рыхлым. Эта теория бвла названа теорией многосечатого роста.
Возникая на ранних стадиях развития того или иного органа, волоски особенно густо располагаются на молодых частях растения. По мере роста листа или другого органа опушение становится более редким, так как в процессе разрастания клеток эпидермиса расстояния между волосками увеличивается. Так, например, молодые нижние листья Verbascum thapsus длиной 3 - 5см имеют сплошной войлочной покров из ветвистых волосков, полностью же сформировавшаяся листовая пластинка опушена значительно слабее.
Жизнь волосков, как правило, недолговечна. Нередко они начинают отмирать с окончанием роста несущего их органа или даже раньше. Однако в других случаях они находятся в жизнедеятельном состоянии значительно дольше. Это чаще наблюдается у железистых трихом, но и простые кроющие волоски могут быть продолжительное время живыми.
И частности, папиллы остаются живыми на протяжении всемго периода жизни несущего их органа. Довольно долго остаются жизнедеятельными и кроющие волоски злаков [5] .
1. 4 Функциональное значение трихом
Функции, выполняемые трихомами, чрезвычайно многообразны. Однако их роль в жизни растении остается далеко не всегда ясной. Наиболее детально изучено функциональное значение специализированных трихомы росянок, с помощью которых насекомоядные растения высасывают питательные вещества из тела насекомого. Издавне и до настоящего времени подвергается глубокому изучению функция корневых волосков, связанная с поглощением элементов минерального питания. Хорошо изучена роль специализированных волосков типа прицепок, с помощью которых происходит распространение семян и плодов [9] . Волоски многих вьющихся и цепляющихся растений, имеющие форму крючков, поддерживают стебель растения. Нередко трихомы выступают в качестве защитных приспособлений. К ним, например, относятся щетинистые волоски бурачниковых и жгучие волоски крапивы.
... продолжениеВы можете абсолютно на бесплатной основе полностью просмотреть эту работу через наше приложение.
- Информатика
- Банковское дело
- Оценка бизнеса
- Бухгалтерское дело
- Валеология
- География
- Геология, Геофизика, Геодезия
- Религия
- Общая история
- Журналистика
- Таможенное дело
- История Казахстана
- Финансы
- Законодательство и Право, Криминалистика
- Маркетинг
- Культурология
- Медицина
- Менеджмент
- Нефть, Газ
- Искуство, музыка
- Педагогика
- Психология
- Страхование
- Налоги
- Политология
- Сертификация, стандартизация
- Социология, Демография
- Статистика
- Туризм
- Физика
- Философия
- Химия
- Делопроизводсто
- Экология, Охрана природы, Природопользование
- Экономика
- Литература
- Биология
- Мясо, молочно, вино-водочные продукты
- Земельный кадастр, Недвижимость
- Математика, Геометрия
- Государственное управление
- Архивное дело
- Полиграфия
- Горное дело
- Языковедение, Филология
- Исторические личности
- Автоматизация, Техника
- Экономическая география
- Международные отношения
- ОБЖ (Основы безопасности жизнедеятельности), Защита труда
